· Ossification enchondrale : c’est l’ossification du squelette cartilagineux embryonnaire. Elle est réalisée par la destruction des cartilages et leur remplacement par des os. L’ensemble des ces os forme l’endosquelette (exemple : l’omoplate). Il est aussi appelé squelette primaire.
· Ossification dermique : les os formés par ossification dermique proviennent du derme et forment le bouclier : l’exosquelette.
Chapitre 3 - Partie 1 :
Le Crâne.
Le crâne est formé par deux parties distinctes :
- Le neurocrâne : c’est la partie qui enveloppe l’encéphale. Il se forme à partir des premiers segments antérieurs. C’est une boite perforée d’orifices qui permettent la communication avec les nerfs et le passage du sang.
- Les capsules sensorielles ou Splanchnocrâne : cette partie est métamérisée. A l’origine, elle est représentée par la succession des arcs branchiaux. Elle entoure les voies aériennes et digestives supérieures.
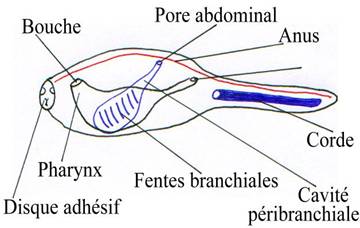
I Les agnathes.
A Le neurocrâne.
C’est une simple boite qui se forme à partir de la lame cartilagineuse située sous le cerveau. Elle se développe en se retournant sur elle-même. Elle englobe latéralement l’encéphale.
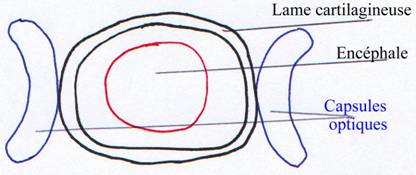
La capsule optique se développe et vient se fixer sur le neurocrâne.

La capsule otique, plus en arrière, sert à l’équilibration et non à l’audition. Elle est disposée orthogonalement. Les canaux sont ciliés et remplis de liquide. Dans ces canaux, se baladent des organites qui vont stimuler une zone ou une autre pour renseigner le cerveau sur la position du corps dans l’espace. Cette capsule s’insère aussi sur le neurocrâne.
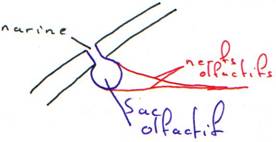
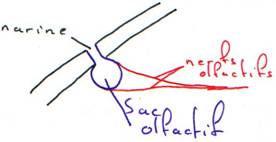
La capsule olfactive (nasale) : les agnates avaient une seule narine médiane (et un troisième œil).
B Le splanchnocrâne.
La bouche est sans mâchoire mais avec une membrane perforée à l’avant par la bouche et latéralement par une série de douze à quinze orifices branchiaux. Chaque poche branchiale est séparée des autres par un arc osseux.
Les arcs branchiaux sont sous le neurocrâne.
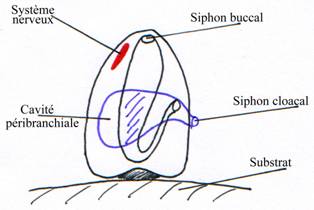
Cas des agnathes actuels (Lamproie).
Les arcs branchiaux ont reculé pour l’adaptation au régime alimentaire.
Le piston lingual est sous le neurocrâne.
Les yeux sont latéraux et réduits à cause du parasitisme.
Les capsules otiques sont à l’arrière du crâne et il n’y a pas de région occipitale.
Le squelette reste cartilagineux.
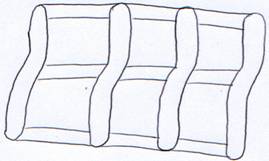
Il ne reste que sept fentes et arcs branchiaux. Les arcs sont non segmentés, reliés entre eux par les travées cartilagineuses. L’ensemble forme la corbeille branchiale.
II Les chondrichtyens (poissons cartilagineux).
A Le neurocrâne.
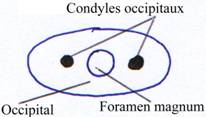
· Il apparaît une région occipitale perforée par le foramen magnum qui laisse passer la moelle épinière.
· Dans la capsule otique apparaît un troisième canal semi-circulaire, disposé perpendiculairement aux deux autres : c’est l’apparition de la perception en 3D.

· La région transorbitale : il y a développement de l’orbite et donc de l’œil.
· La région naso-étnoïdale (museau ou nez) : la capsule nasale a migré à l’avant. Ici, le système est pair : 2 narines.
B Le splanchnocrâne.
Le premier arc branchial disparaît en allant s’intégrer au neurocrâne. Les deuxième et troisième arcs branchiaux vont se céphaliser.
- Le deuxième arc se place sous le neurocrâne pour donner les mâchoires, formées de deux pièces cartilagineuses. La pièce supérieure donne le cartilage palato-ptérygo-carré. La pièce inférieure donne le cartilage de Meckel. Ces deux pièces sont articulées par l’articulation mandibulaire.
- Le troisième arc (l’arc hyoïde, la partie supérieure hyomandibulaire) sert à l’attache des mâchoires sur le neurocrâne è les mâchoires ont une suspension hyostilyque. Le partie ventrale de cet arc passe entre les mâchoires et va soutenir le plancher de la bouche.
Les arcs suivants restent au niveau pharyngien, au nombre de 5, chacun formé de plusieurs pièces. Ils vont soutenir les fentes branchiales (5 paires).
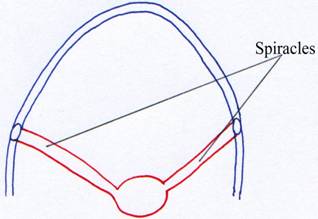
La fente branchiale entre les arcs deux et trois régresse, perd sa fonction respiratoire et subsiste sous la forme d’un canal reliant la bouche (son arrière) au milieu extérieur : c’est l’évent ou spiracle.
III Les ostéichtyens (poissons osseux).
A Le neurocrâne.
On y trouve les quatre régions :
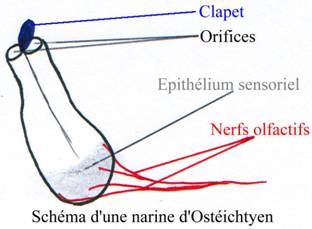
A l’avant on a les narines paires. Chacune possède deux orifices qui conduisent à un sac olfactif tapissé d’un épithélium sensoriel qui transmet l’information à l’encéphale.
Les deux régions intermédiaires ne changent pas.
Au niveau de la région occipitale : la première vertèbre ne s’articule pas sur celle-ci mais s’y imbrique : la tête est donc immobile.
B Le splanchnocrâne.
On assiste à une double ossification du squelette.
· Les arcs mandibulaires : sur la partie supérieure, le cartilage palato-ptérygo-carré s’ossifie en trois points (ossification enchondrale) et donne, le palatin à l’avant, le ptérygoïde au milieu et la carré à l’arrière. Le cartilage de Meckel ne s’ossifie qu’en un point pour donner un os enchondral : l’articulaire.
· L’ossification dermique permet la formation de nouveaux os. A l’avant, on a le pré-maxillaire, puis, le maxillaire. Sur la mâchoire inférieure apparaissent le dentaire et l’angulaire (petit os à l’angle de la mâchoire).
On passe finalement d’une structure à deux pièces à une structure à seize pièces.
Il apparaît une articulation des mâchoires entre le carré (supérieur) et l’articulaire (inférieur). C’est l’articulation reptilienne (elle ne changera que chez les mammifères).
L’arc hyoïde : la partie supérieure s’ossifie pour donner deux os : le hyomandibulaire et le symplectique. Ils suspendent les mâchoires au neurocrâne (suspension hyostylique). La partie inférieure de cet arc s’ossifie en plusieurs pièces sur le plancher de la bouche.
Les cinq arcs branchiaux s’ossifient et gardent leur fonction respiratoire de soutien des fentes branchiales.
L’évent se développe chez l’embryon et disparaît chez l’adulte.
IV Les amphibiens.
Le crâne se modifie à cause de l’adaptation à la vie terrestre selon deux phénomènes : d’une respiration branchiale, on passe à une respiration pulmonaire (régression des fentes branchiales) et on passe du milieu aquatique au milieu terrestre (la transmission des sons est différente).
Il apparaît une structure capable d’enregistrer les variations de pression de l’air : ce sont les organes acoustiques.
Les arcs mandibulaires viennent se souder à la base du neurocrâne : on passe à une suspension autostylique. La partie supérieure de l’arc hyoïde est libérée.
Le hyomandibulaire régresse pour donner un nouvel os : la columelle. Elle se place dans la fente branchiale entre les arcs hyoïde et mandibulaire : c’est l’oreille moyenne.
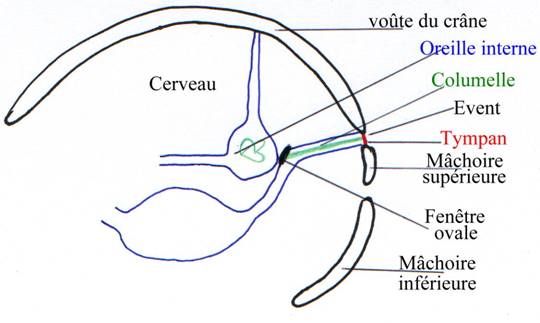
Les vibrations du tympan sont transmises à la columelle puis à la fenêtre ovale. La communication entre la bouche et les oreilles persiste sous la forme des trompes d’Eustache. Elles permettent d’équilibrer la pression de part et d’autre du tympan.
Le reste de l’arc hyoïde (partie ventrale) plus les 5 fentes branchiales se réduisent, régressent, fusionnent pour donner un nouveau système (avec des traces de métamérie) : l’os (ou le complexe) Hyoïde. Dans la plupart des groupes, il ne s’ossifie pas. Il soutien la langue, le larynx, la trachée et la pomme d’Adam. L’arc mandibulaire reste inchangé.
Au niveau du palais apparaît une nouveauté. Le palais est formé par une succession d’os. Le Vomer est à l’avant, suivi du parasphénoïde. è C’est le palais primaire. Chez les grenouilles, les narines percent ce palais et communiquent alors avec la bouche par le choane.
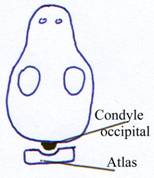
Le crâne s’articule sur la première vertèbre (l’Atlas) et devient mobile.
V Les reptiles.
A partir du moment où le l’arc mandibulaire est soudé au neurocrâne, on ne peut plus distinguer le neurocrâne du splanchnocrâne.
L’arc mandibulaire est inchangé.
Il y a apparition de la mastication. Pour cela, il faut que se développent les muscles de la mastication.
A Les fosses temporales.
Il y a trois types de crânes chez les reptiles.
1 Le crâne anapside.
Progressivement, le nombre de ses os diminue. Le jugal est appelé malaire.
Sur la paroi de la cavité temporale s’insèrent les muscles masticateurs fixés aussi sur la mâchoire inférieure.
2 Le crâne diapside.
La voûte continue se perfore de deux orifices (fosses temporales) qui libèrent les muscles qui peuvent maintenant se développer pour une meilleur fixation et une meilleure insertion.
3 Le crâne synapside.
Les barres (les os qui séparent les fosses temporales) ont disparu : les deux fosses communiquent entre elles. La barre inférieure disparaissant, le quadrato-jugal disparaît aussi. Le carré se trouve ainsi libre et vient alors s’articuler sur le neurocrâne pour permette une grande ouverture de la gueule.
B Le palais secondaire.
A l’avant de la gueule on trouve le pré-maxillaire, le maxillaire. Au milieu se trouvent le Vomer et le parasphénoïde.
Les os latéraux émettent des extensions qui vont recouvrir le palais primaire (les palatins) : on ne voit plus les narines internes.
Entre les deux palais s’est formé le canal naso-palatin qui longe la bouche vers l’arrière et qui débouche au fond de celle-ci par les choanes secondaires.
La respiration avec la bouche pleine est maintenant possible.
VI Les oiseaux.
L’articulation des mâchoires est réalisée entre le carré et l’articulaire.
La suspension du crâne est autostylique. Le crâne est diapside avec disparition de la barre et de l’os post-orbitaire. Réunion avec l’orbite pour former la grande fosse.
Les dents sont remplacées par un bec corné (comme chez la tortue).
La structure de l’oreille est identique.
VII Les mammifères.
Le crâne : il dérive du crâne d’une lignée de reptiles mammaliens (fossiles) de type synapside (une seule paire de fosses temporales inférieures). Ces reptiles sont aussi appelés reptiles synapsides.
Le passage aux mammifères s’est fait en donnant une cavité (orbite + fosse temporale). Le quadrato-jugal disparaît et l’espace laissé libre (à la base) est comblé par une apophyse du jugal détachée du crâne. Celle-ci rejoint une apophyse du squamosal et forment les zygomatiques (les pommettes chez l’homme).
On a un développement de la cavité cérébrale.
Les zygomatiques permettent l’amélioration de la mâchoire. Le palais secondaire est bien formé.
La grande nouveauté est un changement de la structure de l’articulation des mâchoires. Le dentaire se développe, émet une branche qui vient directement s’articuler sur le squamosal (le carré, l’articulaire et l’angulaire se détachent).
La mastication est squamosale.
Les deux dentaires se soudent à l’avant.
Chapitre 3 - Partie 2 :
Le squelette axial.
On distingue deux étapes.
I Le squelette primitif.
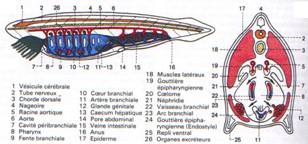
Il est représenté par la corde. Ce sont des cellules turgescentes (haute pression interne) empilées (selon un axe). L’ensemble est retenu dans une gaine formée d’un tissu fibro-élastique.
On la trouve dans tous les embryons de vertébrés et elle persiste chez les cyclostomes et les esturgeons.
La corde est un axe qui ramène le corps à sa position initiale après une courbure.
La corde sera remplacée par le squelette axial secondaire.
II Le squelette secondaire : le rachis.
A Formation du rachis.
Le sclérotome produit des cellules mésodermiques qui migrent vers la partie dorsale où elles entourent le tube nerveux et la corde. Ce mésenchyme donnera une pièce cartilagineuse : la vertèbre.
Particularité : cette vertèbre se forme entre deux métamères successifs.
Chaque vertèbre se forme avec du mésenchyme de deux sclérotomes successifs : la vertèbre est une pièce intermétamérique è on distingue le segment vertébral.
B Evolution de la vertèbre.
La vertèbre la plus primitive est formée de deux pièces dorsales : les basidorsaux qui entourent le tube nerveux et qui vont former l’arc neural.
Les deux pièces ventrales identiques (basiventraux), forment un arc autour de l’aorte : l’arc hémal.
Au niveau de la jonction des basiventraux et des dorsaux, on a la formation de l’apophyse neurale (épineuse) en haut et de l’apophyse hémale en bas.
Au centre, la corde persiste intégralement : on a une vertèbre aspondyle (acentrique).
On trouve ce type de vertèbre chez les agnathes (lamproies) et chez quelques poissons cartilagineux. Chez ces derniers, il apparaît une pièce centrale qui va entourer la corde : le corps vertébral. Il est perforé en son centre par un orifice où passe la corde à apparition des vertèbres classiques.
Chez les poissons cartilagineux, cette structure ne s’ossifie pas contrairement aux poissons osseux.
Dans tous les cas, les vertèbres s’articulent entre elles grâce à la corde.

Chaque vertèbre présente une cavité sur ses faces antérieure et postérieure : ce sont des vertèbres de type amphicoele. La corde est étranglée et se renfle donc dans les espaces intervertébraux. Chaque vertèbre s’articule sur la vertèbre précédente et sur la suivante.
Chez les chondrichtyens, les arcs ne sont pas soudés sur le corps vertébral. Chez les ostéichtyens, tout s’ossifie et les arcs se soudent sur le corps vertébral en donnant une pièce unique. Il existe deux types de vertèbres selon la région du corps :
- la région caudale possèdera des vertèbres typiques, fermées ventralement,
- au niveau du tronc, les arcs hémaux régressent et ne se ferment plus autour de l’aorte (à libération de place).
Il se développent sur les vertèbres des structures d’articulation qui vont remplacer la corde (car elle disparaît à l’état adulte).
A partir des amphibiens, l’arc hémal disparaît totalement (c’est une caractéristique des tétrapodes). Le corps vertébral ne sera plus perforé.
L’arc neural développe deux apophyses latérales : les diapophyses.
Il se développe deux apophyses vers l’avant et deux vers l’arrière : ce sont les zygapophyses. Celles partant vers l’avant prennent appuis sur celles de la vertèbre qui les précèdent.
La corde est remplacée par un coussinet cartilagineux. Celui-ci va se souder au corps vertébral.
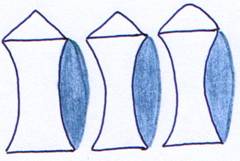
Les vertèbres ont donc leur face postérieure bombée et leur face antérieure creuse : ce sont les vertèbres procoeles.
Remarque : ce sont les vertèbres des anoures et des reptiles actuels.
On a un second cas :
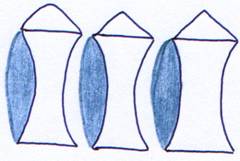
Le cartilage vient se souder à l’avant. Ce sont les vertèbres opistocoeles. On les trouve chez les urodèles.
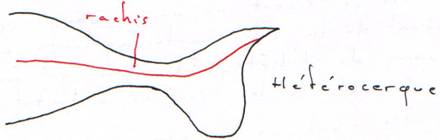
Chez les oiseaux, à cause de la transformation des membres en ailes, le cou est devenu plus allongé et très mobile. Ce sont les vertèbres hétérocoeles. Elles présentent sur chaque face une protubérance et une cavité.
Chez les mammifères, les vertèbres sont biplanes, sans cavité où peuvent se placer les coussinets. Les coussinets deviennent donc également biplans et sont appelés disques intervertébraux.

C La régionalisation.
1 Les poissons osseux.
· De la tête à l’anus, on a la région troncale (avec des vertèbres à arc hémal ouvert).
· De l’anus à la queue, on a la région caudale (vertèbres à arc hémal fermé).
2 Les amphibiens.
Le membre postérieur devient prépondérant (porte la charge maximale). Ce membre va donc se fixer sur la colonne vertébrale par l’intermédiaire d’une ceinture sur les vertèbres sacrées. La région sacrée s’insère entre la région troncale et la région caudale.
La première vertèbre se différencie et ne porte plus de côte : c’est l’atlas.
3 Les reptiles.
Il y a un développement de la région sacrée (au moins deux vertèbres). La partie troncale se subdivise à cause de la formation du cou, d’où l’apparition de la région cervicale. Sur celle-ci, il n’y a plus de côte (elles sont disparues ou très réduites). On a maintenant quatre régions.
4 Les mammifères.
La région sacrée se renforce aussi. La région cervicale, différenciée, ne porte plus de côte. De plus, on a une stabilisation à sept vertèbres cervicales. On trouve deux exceptions à cette règle : le Lamentin et le Paresseux qui peuvent en avoir entre 6 et 9. Maintenant, la longueur du cou dépend de la longueur de chaque pièce. On peut aller de 40 cm pour la girafe à 1,5 cm pour le dauphin.
La région dorsale se fragmente du fait de l’apparition d’une cloison (le diaphragme) dans la cavité abdominale. La partie postérieure est dépourvue de côte : c’est la région lombaire. La région troncale est maintenant la région thoracique.
Chez l’homme, à cause de la bipédie, apparaissent des courbures supplémentaires.
La région caudale est très variable (de 1 à 49 vertèbres) et, est fonction de la longueur de la queue. Chez l’homme, il y a fusion de 3 à 5 vertèbres caudales pour former le coccyx.
5 Les oiseaux.
Leur colonne est divisée en 5 régions. La partie la plus développée est la région cervicale (23 vertèbres cervicales chez le cygne). On assiste à une fusion des vertèbres dans la région postérieure : les dernières lombaires se soudent aux sacrées et aux caudales.
Le premières lombaires et les thoraciques tendent à se souder en un unique os.
C’est une tendance à la rigidification.
D Articulation de la colonne avec le crâne.
C’est l’articulation cranio-vertébrale.
Chez les poissons, il n’y a pas d’articulation : la première vertèbre est imbriquée, la tête est donc immobile.
Chez les amphibiens, il y a différenciation de la première cervicale.
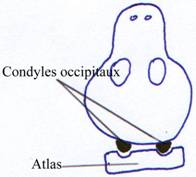
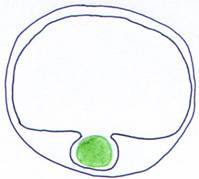
On a maintenant une articulation en deux points qui forme une charnière qui permet le mouvement de bas en haut et de haut en bas de la tête.
Chez les oiseaux et les reptiles, il apparaît une acquisition totale de la mobilité de la tête.

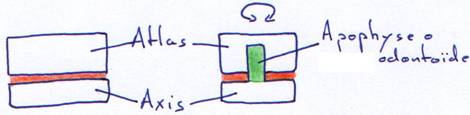
Chez les mammifères, on retourne à une structure à deux condyles latéraux qui servent donc, seulement de charnière. La rotation de la tête se fait entre la première et la deuxième vertèbre cervicale.
III Les côtes.
Ce sont des formations osseuses qui se forment entre les somites par ossification des membranes conjonctives délimitant les métamères. Elles ont donc une structure métamérisée.
On ne trouve pas de côtes chez les agnathes. C’est à partir des poissons qu’elles apparaissent. On en trouve de deux types :
- Les côtes qui bordent la cavité abdominale et s’appuient sur les restes de l’arc hémal : ce sont les côtes monocéphales (ou pleurales ou ventrales).
- Les côtes dorsales. Elles sont placées horizontalement entre les filets des musculatures dorsale et ventrale.
Chez les amphibiens, les côtes sont réduites et ne forment pas de cage thoracique.
Elles sont très développées chez les autres tétrapodes bien qu’il ne reste que les côtes ventrales (les dorsales ayant disparu à partir des amphibiens).
Les côtes bicéphales s’appuient sur le corps vertébral et sur l’apophyse transverse.
Généralement, on a une multiplication des côtes avec la disparition des pattes (y compris sur la région n’en ayant pas d’habitude).
Chez les oiseaux, chaque côte porte une expansion latérale dirigée vers l’arrière : c’est le processus unciné : chaque côte s’appuie sur la suivante pour un renforcement de la cage thoracique.
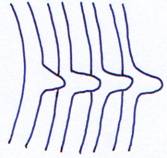
IV Le sternum.
C’est une pièce ventrale métamérisée qui n’existe pas chez les poissons.
Il permet un renforcement du squelette axial en venant se fixer sur les côtes. Il y a formation d’un anneau osseux complet contenant la cage thoracique.
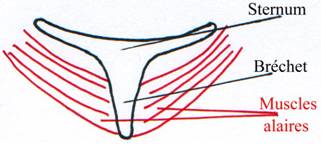
Le sternum disparaît chez les serpents (à cause de la locomotion) et se différencie chez les oiseaux : apparition d’une carène médiane (le bréchet) sur laquelle se développent les muscles alaires.
Chapitre 3 - Partie 3 :
Le squelette des ceintures.
I La ceinture pectorale (ou scapulaire).
A Chez les chondrichtyens.
Elle est très faible, formée de 2 pièces qui s’unissent pour former une cavité d’articulation où s’insère la nageoire pectorale. Ce système est renforcé par l’union médian de ces deux pièces.
La ceinture est sans relation avec le reste du squelette.
Chez certaines espèces, cette ceinture se referme dorsalement ; chez d’autres, elle peut se fixer sur la colonne.
B Chez les ostéichtyens.
On assiste à une multiplication des pièces osseuses. L’ossification enchondrale de ces cartilages donne la scapula et le coracoïde. Ces deux os forment la ceinture primaire où viennent se rajouter d’autres pièces comme le cleithrum (incurvé), le supra-cleithrum et le post-temporal. L’origine de ces trois os est dermique. Ils forment la ceinture secondaire qui suspend la nageoire pectorale sur le crâne (grâce au post-temporal).
La ceinture primaire est réduite alors que la secondaire est prépondérante.
C Chez les tétrapodes.
C’est l’inversion totale.
La ceinture pectorale n’a plus de relation avec le reste du squelette.
La ceinture secondaire régresse pour laisser dominer la primaire.
La ceinture primaire a une structure stable caractéristique. Elle est formée de trois os disposés en étoile qui se rejoignent pour former la cavité glénoïde (cavité d’insertion du membre). La disposition est constante :
- la scapula (omoplate) est dorsale,
- le pro-coracoïde est à l’avant,
- le coracoïde est à l’arrière.
Au cours de l’évolution, un os de la ceinture secondaire, la clavicule, qui apparaît chez certains poissons osseux, vient doubler le pro-coracoïde.
Ce dernier va finir par disparaître pour laisser place à la clavicule.
C’est une ceinture stable qui s’adapte en fonction du mode de locomotion.
1 Les oiseaux.
· Les muscles alaires, insérés sur le bréchet, tirent sur l’humérus. Ils tendent à écraser la cage thoracique. La ceinture est modifiée pour s’adapter à cela. Le coracoïde se renforce et se dispose en « V » sur le sternum pour résister à la contraction des muscles alaires.
· Quand les muscles se contractent, ils tendent à ouvrir cette ceinture : on a donc la soudure des deux clavicules en un os en forme de fourche (« la fourchette »). Cet os a un rôle de maintien de l’écartement constant des deux membres.
· L’omoplate se développe, s’allonge pour assurer un meilleur ancrage de la ceinture dans la musculature en atteignant le bassin (l’omoplate est alors appelée le « sabre »).
2 Les mammifères.
· Au cours du développement embryonnaire, le coracoïde s’atrophie et se soude sur l’omoplate : il donne l’apophyse coracoïde de l’omoplate.
· Pour un meilleur ancrage musculaire, la surface de cette omoplate augmente (par élargissement).
· Il apparaît une crête médiane sur l’omoplate : la crête épineuse. Elle délimite deux fosses épineuses pour une meilleure insertion musculaire. Cette crête s’allonge vers l’avant, dépasse la cavité de l’articulation et forme une apophyse : l’acromion où vient prendre appuis la clavicule.
Remarques :
- Chez l’homme, la clavicule est reliée au sternum.
- La clavicule est bien développée chez les primates et les chauves-souris. Ils ont un membre antérieur très libre (il peut faire des circonvolutions).
- Chez les autres mammifères (les carnivores), le membre tend à se bloquer dans un plan, n’ayant plus que des mouvements pendulaires. Il y a ainsi diminution des risques d’arrachements, d’où, la diminution de la clavicule.
- Chez les vaches et les chevaux, le membre est totalement bloqué ; la clavicule a disparu.
- Selon le type de clavicule, on peut connaître les mouvements du membre.
II La ceinture pelvienne.
A Les poissons.
Cette ceinture est faible et libre dans la musculature. Elle n’a pas de rapport avec la colonne vertébrale. Elle est bâtie sur le principe de la ceinture pectorale des poissons cartilagineux.
Chez certains poissons, elle migre vers l’avant et peut même, chez d’autres, dépasser la ceinture pectorale.
B Les tétrapodes.
Le membre postérieur devient prépondérant (il supporte beaucoup de poids). Cette ceinture vient se fixer sur la colonne vertébrale au niveau des vertèbres sacrées. Elle est exclusivement primaire.
Chaque demi-ceinture est formée de trois os en étoile, formant l’articulation du membre postérieur : la cavité cotyloïde (acétabulum en médecine).
L’ilion : c’est un os dorsal qui se fixe sur les vertèbres sacrées.
L’ischion : c’est un os ventral, dirigé vers l’arrière. Les deux ischions se soudent et donnent la symphyse ischiatique.
Le pubis : c’est un os ventral, dirigé vers l’avant. Les deux pubis se soudent et donnent la symphyse pubienne.
Finalement, on a un bassin compact, en anneau (osseux) autour du corps.
Quelques adaptations :
· Cette ceinture ne régresse qu’avec la disparition des membres postérieurs. On en a l’exemple chez les ophidiens et chez les cétacés. Il reste toutefois quelques rudiments de ceinture chez le boa.
· Les oiseaux ont une attitude bipède avec une colonne horizontale : il y a donc un renforcement caractéristique de cette ceinture pelvienne : les ilions s’allongent, les ischions viennent se réunir avec les ilions en formant une membrane osseuse qui les réunit.
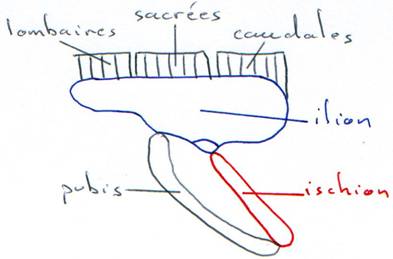
· Le pubis est modifié pour la bipédie : il ne se développe pas vers l’avant mais émet une apophyse vers l’arrière (le post-pubis). On a une « inversion » du pubis qui parfois, se soude sur l’ischion.
Remarque : cette inversion a été retrouvée chez les dinosaures bipèdes. Elle résulte du transfert des muscles qui passent sous la cuisse (c’est une adaptation).
· Chez les oiseaux, il n’y a pas de soudure du pubis et des ischions (ponte des œufs ?!).
· Sous l’action d’hormones, la soudure entre les deux pubis se relâche pour augmenter le diamètre afin de laisser passer le bébé.
· Chez l’Homme, on a un raccourcissement en hauteur du bassin et un basculement par rapport à la ceinture.
Chapitre 3 - Partie 4 :
Le squelette des membres.
I L’ichtyoptérygium.
C’est un simple repli du tégument qui est soutenu par une formation squelettique. On en trouve de deux types :
A Membres impairs.
On trouve la nageoire dorsale et la nageoire anale. Elles ont un rôle dans l’équilibration (stabilisateurs verticaux). La nageoire caudale.
Les poissons nagent par ondulation du corps, de la tête vers la queue : la nageoire caudale permet ainsi d’augmenter la surface qui bat l’eau pour une meilleure propulsion.
Ces nageoires disparaissent au passage à la vie terrestre. Celle qui subsiste le plus longtemps est la nageoire caudale.
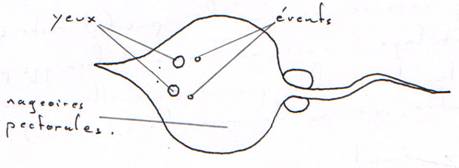
B Les nageoires paires.
Ce sont les nageoires pectorales et pelviennes. Elles ont un rôle de stabilisateurs et de freins grâce à l’orientation mobile à la base.
Au départ, on a une structure complexe.
· Nageoire pluribasale :
La base est large et fonctionne sur le principe des charnières.
· Nageoire monobasale :
On a une pièce qui prend appui sur la ceinture, elle est donc en relation avec la mâchoire. On a alors une grande mobilité (rotation de la nageoire dans toutes les directions de l’espace).
Les nageoires vont acquérir un axe central osseux :
- Nageoire monobasale bisériée : Cette acquisition permet la pénétration de la musculature le long de l’axe (deux séries latérales d’os). Les extrémités membraneuses sont soutenues par les lépidotryctes. Le déplacement sur un substrat solide est maintenant possible.
- Nageoire basale dichotomique : On la trouve chez les fossiles de cœlacanthes. C’est le même principe que chez les bisériée. La pièce basale va donner deux pièces, puis quatre et enfin huit. Le résultat et l’intérêt sont les même que pour les nageoires bisériées.
II Le Chiridium (ou chyroptérygium) = membre pentadactyle.
A Structure.
Le membre prend une fonction locomotrice. Il devient pluriarticulé pour amplifier la force et soulever le corps. On a trois segments.
1 Le stylopode.
Il est formé d’un seul os : l’humérus pour le membre antérieur et le fémur pour le postérieur. Cet os s’articule grâce au coude ou au genou.
2 Le zeugopode.
Il est formé de deux os : le radius et le cubitus pour le membre antérieur, le tibia et le péroné pour le membre postérieur. Ces os s’articulent avec l’autopode grâce au poignet ou à la cheville.
3 L’autopode.
C’est la main ou le pied.
Sa structure est pluriarticulée et très complexe.
- Le basipode (poignet ou cheville) : il est formé de trois rangées d’os nommés en fonction des os du zeugopode. Pour le membre antérieur, on a : le radial, l’intermédiaire et le cubital. Pour le membre postérieur, on a : le péronéal (calcanéum), l’intermédiaire et le tibial (os tragal).
On a ensuite les os centraux (de 0 à 5 os).
Enfin, on a 5 carpiens (pour la main) ou 5 tarsiens (pour le pied).
- Le métapode : Il est représenté par la paume de la main ou par la plante du pied. Ce métapode est composé de 5 métacarpiens ou métatarsiens.
- L’acropode : Il est représenté par les doigts ou par les orteils. Chaque partie est composée d’une succession d’os (les phalanges) articulés entre eux. Les phalanges étaient nombreuses au départ mais ont maintenant une tendance à la réduction. Chez les mammifères,le doigt 1 (le pouce) a deux phalanges, les doigts 2 à 5 ont tous trois phalanges. De l’intérieur vers l’extérieur, on trouve la phalange, la phalangine et la phalangette. Dans le cas du pouce, c’est la phalangine qui disparaît.
Remarque : Le nombre de cinq doigts est un phénomène secondaire. Au début, chez certains amphibiens on trouvait neuf doigts.
La main humaine est assez primitive, ce qui lui laisse une grande possibilité évolutive.
B Disposition des membres par rapport au corps.
On a trois types de membres.
- Le membre horizontal : on ne le trouve que chez les fossiles. Les trois parties du membre sont alignées dans un plan horizontal. Ces membres ne pouvaient donc pas porter le corps (il devait servir à la nage et à la reptation dans des marécages).
- Le membre transversal : c’est le membre des amphibiens et des reptiles. Le stylopode fait un angle de 90 degrés avec la colonne. Le zeugopode s’est relevé à la verticale (les 3 segments sont dans un plan transversal au corps) è ces membres permettent donc de soulever le corps. L’angle zeugo-stylopodique est dirigé vers le bas : il permet la reptation mais avec une ondulation du corps obligatoire. Le poids du corps repose sur les coudes et les genoux.
- Le membre vertical (parasagital) : Les deux premiers segments se redressent. Seul l’autopode reste au sol : on a maintenant des colonnes où repose le poids du corps. Toute la musculature peut-être utilisée pour la locomotion.
· Le membre antérieur : il effectue une rotation de 90 degrés vers l’arrière : l’humérus est toujours vers l’avant mais l’angle zeugo-stylopodique est maintenant dirigé vers l’avant.
· Le membre postérieur : il effectue une rotation vers l’avant, le fémur est toujours vers l’avant mais l’angle zeugo-stylopodique est dirigé vers l’arrière.
Remarque : Les os du zeugopode se croisent pour ramener les mains vers l’avant : c’est la pronation. Celle-ci est en général permanente chez les mammifères.
Chez les primates, la possibilité de décroiser les os s’appelle la supination.
C Adaptation.
Les adaptations touchent surtout au mode de locomotion.
1 Le membre plantigrade (le plus primitif).
La marche se fait sur la plante des pieds, tout l’autopode repose sur le sol (exemple : l’Homme). L’autopode est court et le métapode est bien développé. Cette adaptation permet la marche. Celle-ci se perfectionne chez l’Homme grâce à la bipédie et sert à l’assouplissement de la démarche. On assiste à la création d’une voûte plantaire sous chaque pied. Le pied repose sur trois zones : le talon, le pouce (gros orteil) et le petit orteil.
2 Le membre digitigrade.
L’accélération de la locomotion (trot ou course) se fait par un allongement des membres (plus grandes enjambées) et par une diminution de la surface au sol. Cette diminution se fait par le soulèvement de l’autopode.
Pour cette digitigradie, le basipode et le métapode se redressent ; le talon est haut, au-dessus du sol. Seuls les doigts (acropode) touchent le sol.
3 Le membre onguligrade.
Dans ce type de membre, on parle de digitigradie onguligrade ou d’onguligradie.
On a un redressement de l’acropode. Tout l’autopode se soulève. Seule l’extrémité des doigts touche le sol par la phalangette. C’est la caractéristique des ongulés. De plus, on a la formation de sabots.
Le doigt 1 disparaît toujours et on a au maximum quatre doigts par patte. On a deux lignées distinctes.
a Les mésaxoniens.
L’axe du membre passe par le doigt 3 (ordre des périssodactyles : zèbres, chevaux, tapirs). Ce doigt 3 est toujours plus développé que les autres.
Les tapirs ont quatre doigts à l’avant et trois doigts à l’arrière (le doigt 5 disparaît).
Les chevaux n’ont plus que le doigt 3 avec un sabot. Parallèlement à la disparition des doigts, les pièces osseuses disparaissent : on a une simplification de l’autopode. Il reste des vestiges des métapodes 2 et 4 sous forme de stylets.
b Les paraxoniens.
L’axe du membre passe entre les doigts 3 et 4.
Chez l’hippopotame, les quatre doigts touchent le sol.
Chez les autres espèces, on a une réduction des doigts externes (2 et 5) :
- Le porc a ses doigts 2 et 5 réduits de telle sorte qu’ils ne touchent plus le sol mais forment deux ergots hauts sur la patte.
- Les cervidés : les doigts 2 et 5 forment deux stylets latéraux.
- Les bœufs et moutons : les doigts 2 et 5 ont disparus ainsi que les structures qui les précèdent sur le métapode et sur le basipode.
On a une fusion progressive des métapodes 3 et 4 (è l’os canon) : c’est une caractéristique des paraxoniens (les arctiodactyles).
4 Adaptation au vol.
On trouve cette adaptation chez des reptiles fossiles (ère secondaire).
a Chez les oiseaux.
Pour former l’aile, on a un rééquilibrage en longueur des trois segments. Le cubitus est le plus développé par rapport au radius. On a un allongement de l’autopode, l’humérus diminue.
Cette adaptation se fait aussi par une disparition des doigts. Le 1 est visible par ses vestiges, les 2 et 3 sont bien développés (ils portent les plumes).
Le membre postérieur s’adapte aussi à cette locomotion. Pour supporter le choc de l’atterrissage, le fémur devient horizontal, la cheville se rigidifie par des soudures, l’articulation passe au milieu de l’autopode et donne un os unique qui se soude sur le tibia : le tibiatarse. De plus, le métapode se soude en un os avec le reste du tarse : le tarsométatarse. Seuls les doigts restent libres.
b Chez les mammifères (les chiroptères).
Les doigts deviennent prépondérants (seul le doigt 1 se réduit : il permettra de s’accrocher au sol). Ces doigts vont soutendre une membrane (expansion tégumentaire) : le patagium.
5 Adaptation à la nage.
On trouve cette adaptation chez les mammifères cétacés. Ils ont une dynamique caudale qui assure la propulsion. Cette dynamique entraîne une régression de la musculature latérale : il ne reste qu’une musculature dorso-ventrale en relation avec la nageoire caudale horizontale.
Les membres postérieurs disparaissent.
Les membres antérieurs donnent des palettes natatoires. Tous les os de ces membres s’élargissent et se raccourcissent (dans la longueur) : l’humérus est court et large, le cubitus et le radius sont courts et longs.
On a une simplification du basipode. Les cinq doigts sont hyper-phalangés (jusqu’à 21 phalanges). Toutes les articulations sont bloquées et des cartilages s’accumulent dans ces articulations sauf dans la basale.
6 Adaptation au saut.
Dans ce mode de locomotion, le membre est un ressort à compression. Les trois segments ont une longueur équivalente (allongementde l’autopode). Le système de saut est le même chez les grenouilles, les kangourous, les lièvres…
On a toutefois deux modalités de saut (de réception) :
- Chez la grenouille, sauteur quadrupède, la propulsion est réalisée par les membres postérieurs et la réception est assurée par les membres antérieurs.
- Chez les kangourous, sauteurs bipèdes, propulsion et réception sont assurées par les membres postérieurs.
7 La reptation.
On a une disparition totale des membres.
Chapitre 4 :
L’appareil circulatoire.
I Généralités.
L’appareil circulatoire est formé à partir du mésoderme. Il permet le transport entre les organes, ce qui permet ainsi la croissance de la taille de l’animal. Cet appareil est d’abord lacunaire puis il tend à devenir clos avec la formation des vaisseaux.
A L’apparition de vaisseaux.
Les artères partent du cœur et sont soumises à la pression. Les parois sont fibro-élastiques et très contractiles.
Les veines ont une paroi plus fine et permettent le retour du sang au cœur.
Les jonctions entre les artères et les veines sont assurées par des capillaires qui permettent l’augmentation de la surface d’échange.
Chez les invertébrés, en général, il n’y a pas d’organe propulseur. La circulation sanguine est alors réalisée par les contractions vasculaires.
Chez les vertébrés, il y a apparition d’un cœur unique, structuré, qui assure la propulsion du sang.
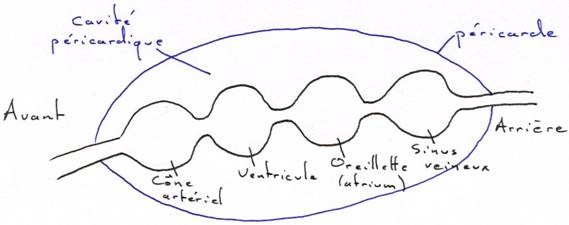
Le cœur est enfermé dans la cavité péricardique. A l’intérieur de cette cavité, le cœur porte l’endocarde dont les parois musculeuses prennent le nom de myocarde.
Chez l’embryon, le cœur est un tube qui, par constrictions circulaires, donne quatre chambres alignées.
Le reflux du sang est empêché par des valvules qui sont développées (surtout) au niveau du cône aortique qui en possède quatre rangées.
Le tube cardiaque s’allonge plus vite que le péricarde et va donc former un repli en « s » du cœur. L’oreillette et le ventricule passent alors en position antérieure.
II L’appareil circulatoire des poissons.
A Système veineux.
Deux veines cardinales parcourent le corps longitudinalement. Chacune est composée d’une veine cardinale antérieure et postérieure. Elles se réunissent pour former le canal de Cuvier qui se jette dans le sinus veineux (système symétrique). Il y arrive aussi une veine qui ramène le sang des viscères (veine sus-hépatique).
B Système aortique ou artériel.
Le ventricule se déplace vers le cône artériel qui régresse (il va former des valvules). L’aorte va ensuite se dilater et donner le bulbe artériel. Le sang quitte le cœur par une artère unique (système impair) qui se subdivise vers chacun des arcs branchiaux : ce sont les arcs aortiques qui sont parallèles et de même nombre que les arcs branchiaux.
Le sang est ensuite pris dorsalement par un vaisseau afférent après chaque arc branchial. Ces vaisseaux vont se réunir pour former dorsalement deux vaisseaux (un de chaque côté) : ce sont les racines dorsales qui dirigent le sang vers l’arrière du corps. Ces deux racines vont se réunir et donner l’aorte dorsale.
A partir des poissons osseux, il ne subsiste que quatre paires d’arcs branchiaux.
Dans un cycle sanguin, le sang ne passe qu’une fois par le cœur.
Le sang n’est propulsé qu’une fois, c’est un système à basse pression.
III L’appareil circulatoire des tétrapodes.
A Généralités.
Quand le cou apparaît, le cœur va reculer en entraînant un raccourcissement de l’aorte ventrale.
Avec le changement de respiration, les poumons n’occupent plus la même place que les branchies : déviation de la circulation vers les poumons, donc, double circulation.
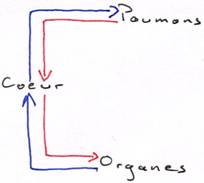
Le sang passe deux fois par cycle par le cœur ; on a donc un système a plus haute pression.
L’évolution va tendre à séparer les deux circulations.
Les veines cardinales des poissons vont être remplacées par les veines caves qui ont le même rôle mais une origine différente.
B Les amphibiens.
1 Le cœur.
Le sinus veineux et le bulbe artériel régressent. Le sinus veineux va se souder à l’oreillette droite et le bulbe aortique au ventricule. L’oreillette présente un cloisonnement grâce à l’apparition d’une membrane médiane. Le ventricule restera unique.
2 Les arcs aortiques.
Chez le têtard, il reste les arcs 3,4,5 et 6. Les arcs de 3 à 5 vont vers les branchies où le sang est hémastosé. L’arc 6 va alimenter la racine aortique dorsale. Une dérivation en part et va alimenter les poumons (qui ne sont pas encore fonctionnels).
On obtient deux artères : l’artère cutanée qui va vers la peau et l’artère pulmonaire qui va aux poumons. Le sang du poumon est ramené à l’oreillette gauche du cœur par la veine pulmonaire.
A la naissance, le poumon est mis sous pression par fermeture de la post-dérivation de l’arc 6 qui donnera le ligament de Botal. L’arc 6 est donc l’arc pulmonaire.
L’arc 5 disparaît.
L’arc 4 (aortique) subsiste et forme les deux racines aortiques (donc l’aorte).
L’arc 3 (carotidien) s’isole et perd sa relation avec le reste de la racine aortique. Il se dirige vers la tête en formant les carotides internes et externes.
L’aorte ventrale va se raccourcir de manière importante.
Le départ de six artères (2×3) va entraîner un mélange des sangs au niveau du ventricule.
Il apparaît toutefois des systèmes contre les mélanges comme, au niveau du bulbe cardiaque, une membrane ondulante qui obture de façon sélective l’un ou l’autre des départs ou par un fonctionnement asynchrone des oreillettes.
Le sang arrive à l’oreillette gauche, passe au fond du ventricule où il est retenu par les cavités multiples creusées dans la paroi du myocarde.
Le sang portant le CO2 arrive par-dessus (stratification des sangs).
Ce sang (veineux) s’en va à la première contraction vers l’arc pulmonaire par la lame spirale.
Quand il y a trop de pression, une valvule bloque l’arc pulmonaire et permet au sang restant de partir vers l’aorte.
B Les reptiles non-crocodiliens.
Chez les amniotes, les arcs 1 et 2 disparaissent très tôt pendant le développement embryonnaire.
1 Le cœur.
Les oreillettes sont cloisonnées, sauf chez les serpents. L’oreillette droite est totalement intégrée avec le sinus veineux : les veines caves se jettent donc directement dans l’oreillette droite.
Le ventricule présente une amorce de cloisonnement. Le bulbe artériel a fusionné avec le ventricule : les arcs partent donc directement du ventricule.
2 Les arcs.
L’aorte ventrale régresse définitivement. Les différents arcs partent donc isolément du ventricule.
L’arc 3 (carotidien) se regroupe et fusionne en un départ unique : le tronc carotidien qui part directement de la crosse aortique : disposition en chandelier.
L’arc 4 est formé de deux crosses aortiques qui partent du ventricule et qui se recourbent vers l’arrière pour fusionner et donner une aorte dorsale unique.
Le tronc pulmonaire (6) fusionne au départ et donne une artère qui se divise en deux en direction de chaque poumon.
On a maintenant une double circulation avec un mélange des sangs au niveau du ventricule (unique) où converge le sang veineux de l’oreillette droite et le sang oxygéné de l’oreillette gauche. La fusion des crosses aortiques permet aussi le mélange des sangs.
C Les reptiles crocodiliens.
1 Le cœur.
Le cloisonnement des ventricules est complet. Les oreillettes sont toujours cloisonnées : on a maintenant deux cœurs latéraux (un droit et un gauche). Il n’y a plus de mélange des sangs au niveau du cœur.
2 Les arcs.
L’arc 3 a une disposition en chandelier sur la crosse aortique droite.
L’arc 6 a la même disposition avec un départ unique depuis le ventricule droit où il reçoit le sang réduit issu des veines caves.
L’arc 4 donne deux crosses aortiques. La crosse gauche part du ventricule droit vers la gauche (charrie le sang veineux). La crosse droite part du ventricule gauche en se courbant vers la droite et transporte le sang oxygéné.
Les deux crosses vont se rejoindre et entraîner un mélange des sangs. A la sortie du cœur, au croisement de ces deux crosses s’établie une communication : le foramen de Panizza. La crosse droite a une pression plus grande que la gauche : le mélange va donc aller plus facilement de la droite vers la gauche, ce qui ne pose pas de réel problème.
D Les oiseaux.
1 Le cœur.
Il est entièrement cloisonné avec un hyper-développement du ventricule gauche (sûrement à cause de l’homéothermie : 41°C). De plus, la consommation d’oxygène des muscles alaires est très élevée. La fréquence cardiaque est comprise entre 300 et 400 battements par minute.
2 Les arcs.
L’arc 4 donne la crosse aortique droite qui prend son départ sur le ventricule gauche et qui véhicule le sang oxygéné.
Le tronc carotidien est branché en dérivation sur cette crosse : il n’y a plus de mélange.
E Les mammifères.
1 Le cœur.
Il présente un cloisonnement complet sauf pendant le développement embryonnaire : on a une relation entre les oreillettes, c’est le trou de Botal qui se rebouche à la naissance.
On a un hyper-développement du ventricule gauche qui envoie le sang dans l’aorte vers tous les organes.
La dissymétrie due à l’hyper-développement oriente la pointe du cœur vers l’axe du corps.
2 Les arcs.
A l’inverse des oiseaux, c’est la crosse aortique droite qui disparaît (pas de mélange des sangs).
La crosse aortique gauche vient du ventricule gauche.
L’arc carotidien se branche sur cette crosse aortique. L’embranchement est différent selon les groupes observés. Les sous-clavières droites s’associent aux carotides droites. Les gauches restent indépendantes.
IV Les modifications de l’appareil circulatoire en fonction des modes de respiration au cours du développement embryonnaire.
Tous les tétrapodes changent de respiration pendant leur développement. Seuls les poissons ne changent pas.
A Les amphibiens.
Chez la larve, les arcs 3, 4 et 5 vont vers les branchies pour assurer les échanges gazeux. L’arc 6 rejoint la racine aortique dorsale. Il y a donc une dérivation sur l’arc 6 qui donne l’artère pulmonaire qui emmènera le sang vers les cellules pulmonaires. Ce sang rejoint l’oreillette gauche par les veines pulmonaires. La base de l’artère pulmonaire présente une constriction qui diminue le diamètre et limite le flux de sang allant vers les poumons.
Changement d’état : le sphincter qui diminue le diamètre de l’artère pulmonaire s’ouvre définitivement d’autant que la relation entre l’arc 6 et l’aorte s’obture totalement en donnant le ligament de Botal.
B Les amniotes.
On a deux cas :
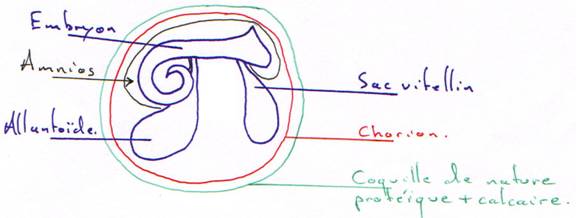
- les ovipares : leur respiration embryonnaire est choro-allanthoïdienne (diffusion d’air à travers la membrane vers l’appareil circulatoire quand il est fonctionnel)
- les placentaires : leur respiration est assurée par les poumons de la mère. Le sang maternel oxygéné sera véhiculé vers l’embryon.
Dans les deux cas, les poumons se développent mais ne doivent recevoir de sang que pour la nutrition : on a besoin d’une communication de l’arc 6 avec l’aorte dorsale et d’une diminution du diamètre de l’artère pulmonaire.
Chez les mammifères, le trou de Botal permet le passage du sang entre les deux oreillettes.
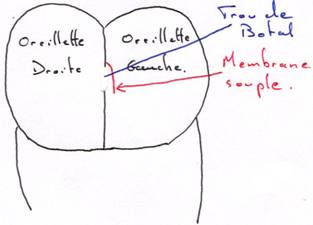
Le trou et le futur ligament de Botal dérivent 92,5% du sang devant aller vers les poumons.
Un shunt est la partie déviant le sang vers l’aorte dorsale.
A la naissance, la relation de l’arc 6 avec l’aorte s’obture par un resserrement du vaisseau dont les parois se soudent (entre deux et sept semaines). On a alors une arrivée brutale de sang dans l’oreillette gauche. La membrane souple va être plaquée sur le trou de Botal. Elle va ensuite se souder et obturer définitivement le passage.
Chapitre 5 :
Le système nerveux.
I Généralités.
C’est un véhicule de l’information sensorielle et motrice vers les effecteurs. Il traite l’information du système nerveux central.
A Le système nerveux central.
Il est protégé par le squelette. Il comprend :
- L’encéphale situé au niveau du crâne qui est prolongé par
- La moelle épinière qui est protégée par le canal rachidien formé par la succession des arcs neuraux des vertèbres.
Le cerveau est composé de ventricules et la moelle épinière est parcourue par le canal de l’épendyme. Ce système nerveux central est formé de cellules nerveuses (neurones) qui, par les dendrites amènent l’information au corps cellulaire et par les axones qui exportent les informations depuis le corps cellulaire.
Les corps cellulaires ont une couleur grise et sont regroupés pour former la substance grise.
Les axones sont recouverts par une gaine de myéline, blanche (sauf chez les cyclostomes). Les axones sont regroupés pour former la substance blanche.
Les cellules gliales vont former la neuroglée.
Les cellules de l’épendyme vont tapisser les cavités internes mais surtout le canal de l’épendyme. Elles constituent une barrière perméable qui permet les échanges entre le liquide céphalo-rachidien et les tissus du cerveau.
B Le système nerveux périphérique.
Il est subdivisé en fonction de l’origine des nerfs. On a :
- Les nerfs crâniens dont le nombre est compris entre 10 et 12 selon le groupe de vertébrés.
- Les nerfs rachidiens qui partent de la moelle épinière entre les vertèbres.
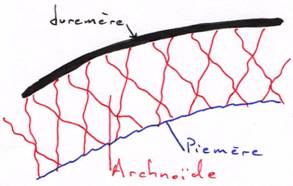
C Les méninges.
Ce sont des structures de protection qui entourent le système nerveux central. On en trouve une seule chez les poissons, deux chez les amphibiens et trois chez les amniotes.
La plus externe est la dure mère (plaquée contre les os) ; au milieu, on a l’arachnoïde qui est une structure lâche en toile d’araignée ; la plus interne est la pie mère (molle).
II Le système nerveux central.
A Origine et formation.
Le tube nerveux se dilate au niveau du crâne pour former une vésicule cérébrale. Elle subit trois constrictions et donne trois chambres successives qui vont correspondre aux trois capsules sensorielles :
- 1 : le proencéphale.
- 2 : le mésencéphale.
- 3 : le rhombencéphale.
Les vésicules 1 et 3 vont subir une nouvelle constriction et donner ainsi 5 vésicules :
- télencéphale
- diencéphale
- mésencéphale
- métencéphale
- myélencéphale.
Primitivement, on a une dilation du canal central pour former les ventricules.
B Anatomie comparée du télencéphale.
1 Généralités.
La prolifération des parois latérales donne deux lobes latéraux (les hémisphères cérébraux). La partie centrale (le télencéphale primaire) donne les hémisphères cérébraux (télencéphale secondaire). Cette étape est la dernière évolution de l’encéphale des cyclostomes.
On a ensuite prolifération des hémisphères vers l’avant (pour les poissons et les urodèles) puis vers l’arrière (pour les anoures et les amniotes).
Le télencéphale secondaire devient dominant et donne deux ventricules qui vont communiquer avec l’extérieur par deux orifices : les trous de Monro. Cette partie avant des ventricules pousse vers la capsule olfactive avec laquelle elle entre en relation : les structures initiales olfactives donnent ainsi les lobes olfactifs.
Les ventricules vont croître vers le haut en se recourbant à l’intérieur et en se rejoignant sur la ligne médiane : on a maintenant une structure inversée du cerveau.
La paroi supérieure de l’hémisphère est formée par le pallium qui contient la substance grise. On distingue :
- L’archipallium. Il est le plus près de la ligne médiane.
- Le paléopallium. Il est le plus latéral.
Le plancher est formé par les corps striés. Au cours de l’évolution se rajoutera le néopallium entre les deux précédentes parties du pallium.
2 Les sélaciens.
Ils ne présentent pas de séparation entre les ventricules : cavité unique.
Le paléopallium est un relais olfactif qui envoie l’information vers le diencéphale (vers le thalamus dorsal). Il reçoit l’information visuelle venant des lobes visuels optiques du diencéphale. Les messages moteurs sont pris en charge par le thalamus ventral.
3 Les amphibiens.
Il y a séparation des deux ventricules par la région septale.
L’information olfactive va vers le paléopallium puis vers l’archipallium et au corps strié.
Les informations visuelles vont au corps strié par le thalamus dorsal.
Le télencéphale récupère toutes les informations sensorielles.
Les informations motrices partent par le thalamus ventral.
4 Les crocodiliens.
Les hémisphères cérébraux augmentent et les bulbes olfactifs s’allongent.
L’encéphale est marqué par un développement de l’archipallium et par le paléopallium latéralement : ils deviennent recouvrant. Les corps striés se retrouvent à l’intérieur où ils s’hyper-développent.
Le télencéphale est marqué par une inversion de la substance grise qui passe à la périphérie.
Il apparaît au niveau du pallium une structure nouvelle : le néopallium. Il s’intercale entre l’archipallium et le paléopallium. On a ainsi une augmentation du développement périphérique du pallium (corps striés à l’intérieur).
Les influx sensoriels visuels sont acheminés au paléopallium et sont transmis à l’archipallium puis aux corps striés qui reçoivent les autres informations sensorielles par le thalamus dorsal.
5 Les oiseaux.
Les corps striés prennent leur maximum d’importance et de développement. La structure du pallium est peu développée. Les hémisphères cérébraux sont hyper-développés (ne concernent que les corps striés).
6 Les mammifères.
Les hémisphères cérébraux présentent leur développement le plus important. Cette croissance résulte de l’hyper-développement du néopallium qui entraîne le rejet de l’archipallium au centre, entre les deux hémisphères. Ces phénomènes entraînent la formation de l’hippocampe.
Le paléopallium est rejeté latéralement et ventralement.
Le néopallium se développe et recouvre vers l’arrière le mésencéphale et le diencéphale.
Toutes les informations sensorielles sont transmises au néopallium qui devient le centre de coordination sensorielle. Il prend aussi le contrôle des informations motrices. C’est maintenant une structure essentielle.
a Les mammifères inférieurs.
La paroi du télencéphale reste lisse : on parle de mammifères lissencéphales. Ce groupe comprend les rongeurs, les marsupiaux, les monotrèmes et les insectivores.
b Les mammifères supérieurs.
Ils sont caractérisés par une nouvelle augmentation de la surface du néopallium (ou néocortex). L’accroissement est réalisé par un plissement de la surface qui donne des circonvolutions profondes : c’est la paroi externe circonvolutionnée des mammifères gyrencéphales.
Au niveau du néocortex, il y a différenciation d’aires bien définies (sensorielles, d’association, motrices) (cf. cours de physiologie animale SVI 04).
C Anatomie comparée du diencéphale.
Le diencéphale se situe à l’arrière du télencéphale.
Il est occupé par une cavité : le troisième ventricule.
Sa paroi dorsale est mince, dépourvue de cellules nerveuses. Une couche s’associe à la pie mère et forme une paroi perméable : la toile choroïdienne. C’est une zone richement vascularisée qui assure les échanges entre le sang et le liquide céphalo-rachidien.
Les parois latérales forment le thalamus qui émet un bourgeon dorsal : l’épithalamus qui est très développé chez les vertébrés inférieurs. Il vient même au contact d’un orifice de la paroi crânienne en donnant le troisième œil ou œil pinéal qui est un organe photosensible et thermosensible. On retrouve cet organe particulier chez un rhynchocéphale, Hattéria, qui est un gros lézard des îles des côtes d’Australie.
L’épithalamus comprend l’épiphyse qui a subsisté malgré la régression de l’œil pinéal. Elle a gardé une sensibilité aux variations de lumière et de température. Elle sécrète la mélatonine pour les cycles saisonniers (reproduction …).
La paroi ventrale développe une glande, l’hypophyse, qui a un rôle endocrinien essentiel. Cette glande est fixée par un pédoncule. Elle est logée dans une cavité de l’os et reliée par l’infontébulum.
Ce diencéphale voit ventralement, pendant son développement embryonnaire, deux expansions se dirigeant vers les capsules optiques qui donneront les nerfs optiques et la rétine. Avant leur entrée dans le diencéphale, on a la mise en place d’un chiasma optique (croisement des fibres optiques).
D Anatomie comparée du mésencéphale.
Chez les anamniotes, la cavité intérieure est dilatée et donne un ventricule.
Chez les amniotes, cette cavité donne un canal étroit : l’aqueduc de Sylvius.
Le mésencéphale est occupé dorsalement par les lobes optiques où arrivent les deux nerfs optiques : c’est un centre de relais dont le développement est en relation avec le développement de la vision.
Chez les anamniotes, on obtient les tubercules bijumeaux.
Chez les amniotes, on aura les tubercules quadrijumeaux. Les deux tubercules antérieurs servent de relais optique et les deux postérieurs servent de relais otique et auditif.
E Anatomie comparée du métencéphale (le cervelet).
La prolifération des parois latérales et surtout dorsales donne une excroissance qui est un centre de coordination motrice réflexe. Elle assure le maintien de la posture. Son développement est fonction de la locomotion.
L’archicérébellum donne les parties latérales et le paléocérébellum donne la parie médiane (le vermis) et chez les mammifères, il produit en bourgeonnant une nouvelle partie de deux extensions latérales : les hémisphères cérébelleux ou néocérébellum.
Les stimuli sensoriels sont canalisés par le néocortex puis transmis au cervelet qui acquiert la fonction de centre de la mobilité volontaire.
F Anatomie comparée du myélencéphale (bulbe rachidien).
Le myélencéphale renferme le quatrième ventricule.
Sa paroi dorsale est une toile choroïdienne. Le plancher et les parois latérales forment le centre de voies réflexes (réflexes primaires). Ce centre contrôle : le rythme cardiaque, les mouvements respiratoires, la déglutition, la toux, les sécrétions salivaires, …
G Anatomie comparée de la moelle épinière.
La moelle épinière est dans l’arc neural des vertèbres.
Primitivement elle va jusqu’à l’extrémité de la queue. Avec la régression de la queue et de la musculature latérale, elle se raccourcie et n’atteint plus la dernière vertèbre.
Elle est, comme l’encéphale, entourée par les méninges.
Chez les lamproies, elle est sous la forme d’une lame.

Chez les gnathostomes, la moelle devient circulaire. La substance grise est toujours interne. Les cornes latérales dorsales renferment les corps cellulaires des neurones sensoriels et les cornes latérales ventrales renferment les corps cellulaires des motoneurones.
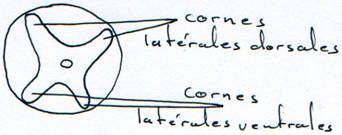
Cette moelle épinière a deux fonctions essentielles : véhiculer les informations vers cerveau ou depuis le cerveau et servir de centre d’arcs réflexes.
III Le système nerveux périphérique.
A Les nerfs crâniens.
Ces nerfs témoignent de la segmentation primitive du cerveau.
On trouve dix paires de nerfs crâniens chez les anamniotes et douze chez les amniotes.
- Les nerfs sensoriels :
· 1 : nerf olfactif
· 2 :nerf optique
· 8 : nerf auditif
- Les nerfs moteurs :
· 3 : le nerf oculaire commun, il innerve quatre muscles moteurs des yeux.
· 4 : le nerf pathétique, il innerve les muscles obliques supérieurs des yeux.
· 6 : le nerf oculaire externe, il innerve les muscles externes des yeux.
- Les nerfs mixtes (sensoriels et moteurs) :
· 5 : le nerf trijumeau
· 7 : le nerf facial, il innerve la peau de la tête, les glandes lacrymales et salivaires ainsi que la partie avant de la langue.
· 9 : le nerf glosso-pharyngien, il innerve le reste de la langue et les muscles voisins des ex-fentes branchiales.
· 10 : le nerf vague ou pneumogastrique, sa branche principale va aller innerver le cœur, le tube digestif et en particulier l’estomac.
· 11 : le nerf spinal. Il est à dominance motrice et innerve les muscles du cou (jusqu’au sternum).
· 12 : le nerf hypoglosse. C’est un nerf principalement moteur qui innerve la langue.
B Les nerfs rachidiens.
Ces nerfs sont issus de la moelle épinière et possèdent deux racines.
L’information sensorielle arrive aux corps cellulaires situés dans le ganglion spinal. Ils transmettent cette information aux autres neurones par les dendrites.
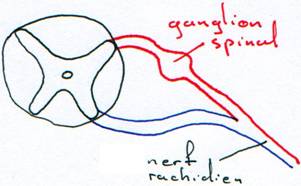
Le nombre de nerfs rachidiens est fonction du nombre de métamères. Il est très élevé chez les poissons ; on en trouve une dizaine chez les amphibiens et trente et une paires chez l’Homme.
Chapitre 6 :
L’appareil urogénital.
Il est composé de deux appareils qui entretiennent des rapports étroits surtout chez le mâle. Les deux parties sont d’origine mésodermique. Ils se forment à partir de tubes issus du cœlome : ce sont les cœlomoductes.
I L’appareil excréteur urinaire.
Il permet la filtration du milieu.
Les éléments de base qui le composent sont les néphrons (cœlomoductes). On a une paire de néphrons par métamères. Ils sont comparables aux néphridies.
A Formation des divers types de reins.
La néphrogenèse ne se fait pas de façon régulière et continue ni dans l’espace ni dans le temps : on a plusieurs types de reins qui se succèdent au cours du temps et dans l’espace.
Il se forme ainsi le premier rein primitif (pronéphros) juste à l’arrière du cou. Il lui succède le mésonéphros dans la région médiane du corps. Chez les vertébrés supérieurs, ce mésonéphros est remplacé par un métanéphros (postérieur).
1 Le pronéphros.
Ce rein n’est fonctionnel que chez quelques poissons à l’état adulte. Ils sont toujours présents à l’état larvaire des anamniotes.
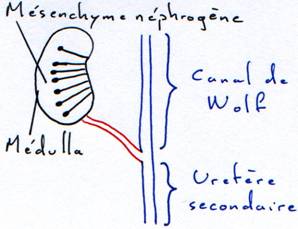
a La formation du canal de Wolf (ou uretère primaire).
A partir d’une ébauche de mésenchyme antérieur dorsal se forme un canal qui progresse vers l’arrière dorsalement jusqu’au cloaque où il va déboucher. La formation de ce canal précède la mise en place des néphrons.
b La formation des néphrons.
La pièce intermédiaire forme un canal qui part du cœlome qui rejoint et se branche sur le canal de Wolf de manière métamérique.
En même temps, le cœlomoducte émet une ramification vers l’aorte dorsale qui en émet aussi une vers lui. Cette ramification se différencie en une capsule externe appelée capsule de Bowman.
Au point de rencontre, le vaisseau se pelotonne et donne une zone dont la surface d’échange va augmenter.
La communication initiale avec le cœlome s’obture. Le tube s’allonge démesurément et le néphron est formé pour assurer la fonction épuratrice du sang.
Le rein est constitué par une succession de néphrons. A l’origine métamérique, ils pourront se différencier par bourgeonnement. Le pronéphros comporte peu de néphrons.
On en trouve 5 chez les lamproies, 2 à 5 chez les amphibiens, 10 chez le poulet et 8 chez l’Homme.
c Le canal de Müller.
Dorsalement, à l’arrière du cou, la cavité cœlomique donne une expansion vers l’extérieur. Ce canal débute par un entonnoir qui prolifère vers l’arrière parallèlement au canal de Wolf.
Ce canal est le gonoducte : le tractus génital femelle.
2 Le mésonéphros.
Il se forme dans la partie médiane de la cavité abdominale. Il remplace le pronéphros chez les adultes anamniotes (poissons et amphibiens).
On y trouve une centaine de néphrons qui débouchent dans le canal de Wolf qui reste l’uretère primaire. Il peut y avoir une multiplication secondaire des néphrons (chez les chondrichtyens et les anoures). Quand tout le tissu mésenchymateux a été utilisé, la multiplication se fait par bourgeonnement des néphrons déjà existant (chez les téléostéens et les urodèles).
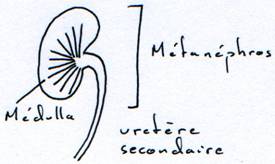
3 Le métanéphros.
Ce métanéphros remplace le mésonéphros. Pro et mésonéphros se succèdent au cours du développement embryonnaire et sont remplacés par le métanéphros dans le « creux des reins » (région lombo-sacrée).
Ce dernier rein se forme à partir d’une masse de mésenchyme indifférencié et d’un bourgeonnement du canal de Wolf. Il se différencie alors les néphrons (è mésenchyme néphrogène).
Dans un second temps, il y a multiplication secondaire des néphrons par bourgeonnement. On a jusqu’à un million de néphrons chez les mammifères.
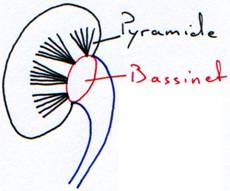
Dans le métanéphros a lieu une zonation où les capsules de Bowman sont regroupées à la périphérie pour donner le cortex. Les néphrons et les tubes se disposent en parallèle et donnent les pyramides de Malpighi.
4 Les différents types de métanéphros.
· Le rein unipyramidaire.
On le trouve chez les primates (sauf l’Homme), les monotrèmes, les marsupiaux et les rongeurs.
· Les reins pluropyramidaires.
- Le rein pluripyramidaire lisse. On le trouve chez l’Homme.
- Le rein pluripyramidaire lobulé. Les pyramides tendent à s’isoler. On trouve ce rein chez les bovidés.

- Le rein pluripyramidaire en grappes. Les pyramides sont séparées. On retrouve ce type de rein chez les cétacés et chez les ours.

B Les structures annexes.
1 La vessie.
Chez de nombreux vertébrés, la vessie est l’organe de stockage de l’urine. Elle est formée par la dilatation des uretères. Chaque uretère débouche isolément dans la vessie qui est extensible.
De cette vessie part l’urètre.
Chez les anoures, la vessie est une différenciation du cloaque. Il émet une dilatation latérale et extensible. Les uretères débouchent dans le cloaque.
2 Le cloaque.
Le cloaque est un carrefour où se déversent le rectum, la vessie et les deux conduits génitaux (quand ils sont séparés). Ces trois appareils ont une partie commune : un orifice unique.
Cette disposition est toujours vraie chez les monotrèmes.
Chez les autres mammifères, il y a formation d’une cloison verticale qui divise le cloaque en deux grandes cavités : une pour le tube digestif (anus, à l’arrière) et une pour le sinus urogénital.
Chez les femelles, le sinus urogénital est séparé par un cloisonnement. On obtient alors trois orifices distincts.
II L’appareil génital.
Les vertébrés sont gonochoriques avec une reproduction sexuée.
Exceptions : - chez les poissons, il peut y avoir un changement de sexe pendant la vie de l’animal. – chez certaines espèces de lézards, on a des cas de parthénogenèse (reproduction de la femelle sans le mâle).
- Dans les groupes inférieurs, la reproduction est externe, sans accouplement.
- La fécondation reste externe mais avec accouplement (crapaud, grenouille). Les mâles fécondent les œufs au fur et à mesure qu’ils sont pondus.
- La fécondation devient interne grâce à des organes sexuels différenciés (chez les amniotes et les chondrichtyens).
è La reproduction devient de plus en plus efficace, avec limitation des pertes, diminution des œufs produits. On a aussi l’apparition des soins aux jeunes.
A La formation des gonades.
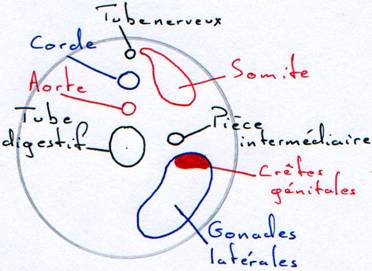
Les crêtes génitales pendent dans la cavité générale par un mésenchyme. Des cellules endodermiques viennent coloniser cette crête. Elles donneront les gamètes.

Parallèlement, des restes de mésenchyme ayant servi à la formation des reins pénètrent dans la crête pour former la partie interne ou medulla.
Jusque là, le développement est commun à tous les vertébrés des deux sexes : on parle de stade indifférencié.
1 Pour le mâle.
Le cortex régresse. La medulla se développe. Elle se creuse de tubes (séminifères). Qui convergent et entre en relation avec des néphrons dont le glomérule ne se développe pas.
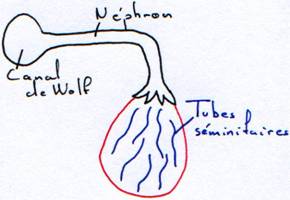
On a maintenant une relation entre les appareils urinaire et génital. Le canal de Wolf acquiert une nouvelle fonction : c’est un canal uro-génital.
2 Pour la femelle.
On a une hypertrophie du cortex et une disparition de la medulla. Les cellules vont se différencier en gamètes : les ovaires restent totalement distincts de l’appareil excréteur.
Les ovules sont excrétés dans la cavité abdominale puis captés par le canal de Müller et transmis aux trompes de Fallope.
B Anatomie comparée des voies génitales.
1 Les agnathes.
On n’a pas de développement de conduits génitaux. Les gamètes mâles ou femelles sont déversés dans la cavité abdominale puis libérés dans le milieu extérieur par un pore situé près de l’anus.
2 L’appareil femelle :
· Les anoures.
Le canal de Wolf est toujours primaire (mésonéphros) ; les voies génitales sont séparées des autres voies. Le canal de Müller est distinct de celui de Wolf : c’est un oviducte.
· Les mammifères.
L’urine du métanéphros est récupérée par les uretères secondaires : le canal de Wolf régresse.
Les ovules pondus sont récupérés par le canal de Müller. Avec la placentation, la paroi de ce canal devient musculaire, se différencie pour former un utérus.
A l’origine, chaque oviducte est touché par cette transformation.


L’utérus bicorne est le plus répandu. L’utérus simplexe est présent chez les primates, les fourmiliers et les édentés.
3 L’appareil mâle.
· Les téléostéens.
Le mésonéphros est collecté par le canal de Wolf. L’appareil génital est indépendant de l’appareil urinaire. On assiste à la formation de deux spermiductes qui se rejoignent un peu avant le pore génital mâle.
· Les anoures.
L’appareil excréteur est un mésonéphros collecté par le canal de Wolf. Pendant le développement embryonnaire, certains néphrons se branchent sur les tubes séminifères : le canal de Wolf devient un uro-spermiducte. Le canal de Müller disparaît : il est exclusivement femelle.
· Les mammifères.
On assiste à la dégénérescence du canal de Müller.
Les testicules sont drainés par des néphrons modifiés et se déversent dans le canal de Wolf.
L’apparition du métanéphros met en place les uretères secondaires.
è Le canal de Wolf devient exclusivement génital : on parle de spermiducte. On a une séparation complète des deux appareils.
Le canal de Wolf se pelotonne la surface du testicule pour former l’épididyme.
Sommaire du cours de Biologie Animale BGU04.
|
Chapitre 1 : Les Protozoaires.
I Le règne des protistes.
II Morphologie et structure des protozoaires.
III Classification.
IV Biologie des protozoaires.
V Association avec d’autres organismes.
VI Les formes coloniales.
|
Chapitre 2 : Les Diploblastiques.
I Les spongiaires.
II Les cnidaires.
III Les cténaires (= les Cténophores).
|
Chapitre 3 : Les Métazoaires Triploblastiques.
I Les Acœlomates.
II Les Pseudocœlomates.
|
|
Chapitre 4-1 : Les Métazoaires Triploblastiques Cœlomates.
L’embranchement des annélides.
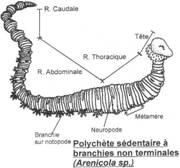
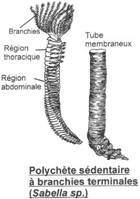
I La classe des Polychètes.
II La classe de clitellates.
III les Pogonophores.
|
Chapitre 4-2 : Les Métazoaires Triploblastiques Cœlomates.
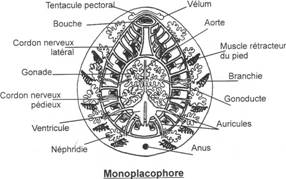
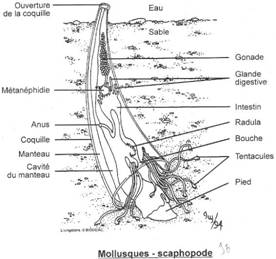
L’embranchement des Mollusques.
I Les caractères communs.
II Etude de classes.
|
Chapitre 5-1 : Les Arthropodes, Généralités.
I Caractères généraux.
II La diversité des arthropodes.
III Les Pararthropodes, groupe associé aux arthropodes.
IV Signification fonctionnelle de la métamérie.
V La céphalisation.
|
|
Chapitre 5-2 : La locomotion chez les Arthropodes.
I L’exosquelette et l’appendice locomoteur.
II La locomotion chez les crustacés.
III La locomotion en milieu terrestre.
|
Chapitre 5-3 : La respiration chez les Arthropodes.
I Les arthropodes aquatiques stricts.
II La vie de la zone de balancement des marées.
III Les arthropodes terrestres.
IV Le retour au milieu aquatique.
|
Chapitre 5-4 : Le Développement Embryonnaire des Arthropodes.
I Généralités.
II Le développement post-embryonnaire des crustacés.
III Développement post-embryonnaire des insectes.
|
|
Chapitre 5-5 : Prédation et Défense chez les Arthropodes.
I La Prédation.
II La réponse des proies : défense contre les prédateurs.
|
Chapitre 6 : L’embranchement des Cordés.
I Généralités.
II Les Agnathes.
III Les gnathostomes.
|
Chapitre 7 : L’Hominisation.
Généralités.
I L’Hominisation.
II L’origine de l’Homme moderne.
III Le cas du peuplement de l’Europe et des Néandertaliens.
IV De qui descendons-nous ?
|
Chapitre 1 :
Les Protozoaires.
Les protozoaires furent observés pour la première fois il y a 300 ans. Ceux sont des unicellulaires, mobiles au moins à un stade de leur développement. Aujourd’hui, ils sont placés dans le règne des protistes.
I Le règne des protistes.
Les protistes sont des unicellulaires et la structure d’une amibe, par exemple, est comparable à n’importer quelle cellule eucaryote : ce sont des organismes autonomes assurant toutes leurs fonctions vitales è ce sont des cellules totipotentes.
Par conséquent, une cellule protiste n’est pas comparable à une cellule de métazoaire mais à un métazoaire en intégralité.
A Protozoaires = Animaux ?
Les unicellulaires autotrophes sont placés parmi les végétaux alors que les unicellulaires hétérotrophes sont rapprochés des animaux.
On peut donc distinguer :
- Les protophytes (affinité végétale) : ils ont un pigment pour la photosynthèse qui assure l’autotrophie. Ils possèdent aussi des constituants des cellules végétales comme l’amidon et la cellulose.
- Les protozoaires : Ils doivent se procurer les substances vitales dans l’environnement. Ce sont les animaux les plus simples.
- Les formes intermédiaires. Exemple : Euglena. Euglena possède des chloroplastes mais si elle est élevée à l’obscurité, elle devient un hétérotrophe irréversible.
B Distribution des protozoaires et importance écologique.
Malgré la simplicité de leur organisation, la structure protozoaire est réussie car la vie protozoaire est présente sous tous les climats et dans tous les habitats. On peut les trouver :
- A l’état libre (en milieu aqueux ou humide).
- Comme parasite (maladie).
- Comme symbiote.
La modification d’un plan structural de base, en vue de rendre les protozoaires capables d’occuper tous les habitats et de nombreux modes de vie est appelée radiation adaptative.
Cette radiation adaptative permet de réduire la compétition entre des animaux semblables à l’origine, ce qui permet l’accroissement de la diversité.
II Morphologie et structure des protozoaires.
A Taille.
Les protozoaires ont une taille comprise entre 1 et 600µm. Les plus petits sont les sporozoaires ainsi que certains parasites intracellulaires. Les plus grands sont les amibes qui peuvent atteindre jusqu’à 5mm.
B Structure.
Les protozoaires possèdent tous les constituants classiques de la cellule eucaryote (organites spécifiques) :
- Membrane lipoprotéique mince : plasmalemme.
- Membrane lipoprotéique parfois doublée d’une enveloppe superficielle. Cette membrane a un rôle de protection contre les agressions et la déshydratation. Lorsque cette dernière est bien développée, on peut trouver une membrane cellulosique, calcaire, siliceuse. On général, on parme de test, de coque, de lorica, de loge…
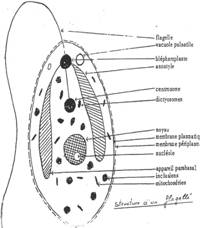
- L’appareil de Golgi (synthèse de membrane). On trouve, à ce niveau, des différences. On observe des empilements de saccules qui forment les dictyosomes. Chez les flagellés, on trouve des dictyosomes très volumineux (ou appareil parabasal) qui ont un rôle dans la sécrétion et l’emballage.
- Le noyau. Chez les protozoaires, le noyau est souvent plurinucléé mais seulement pendant un état transitoire (division du cytoplasme en autant d’individus qu’il y a de noyaux). On trouve toutefois des protozoaires avec constamment deux noyaux : les ciliés (exemple : paramécies) qui possèdent un macronucléus et un micronucléus.
- Les cils et flagelles. Ils ont la même structure chez les protozoaires et les métazoaires (spermatozoïdes). Les cils sont courts et nombreux (5 à 15µm) ; les flagelles sont plus rares et longs (150 à 200µm).
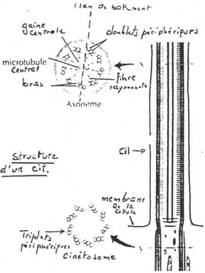
- Le cytosquelette. Il est très développé et constitué par des microfilaments ou des microtubules. Les microfilaments sont constitués d’actine (protéine) et jouent un rôle dans les mouvements (contractions) de la cellule. Parfois, la cellule renferme, le long de son plus grand axe, une structure rigide, « l’axostyle » ou baguette qui est un faisceau de microtubules.
- Les trichocystes. On les trouve chez les ciliés, à la périphérie du cytoplasme. Ce sont des dispositifs de défense et d’attaque. Ce sont des petits dards gorgés de toxine. Ils jaillissent à l’extrémité d’un petit filament pour tuer ou paralyser les proies.
III Classification.
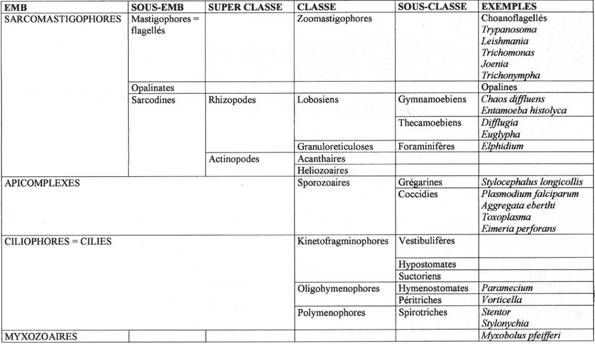
La classification des protozoaires a subit de nombreux remaniements ces dernières années. La principale discrimination se fait en fonction de l’appareil locomoteur.
On trouve quatre embranchements.
A Les Sarcomastigophores.
1 Les Flagellés.
Ils réalisent leurs déplacements grâce à des flagelles. Au cours du cycle, il n’y a pas de spore. La reproduction sexuée est rare.
2 Les Rhizopodes.
Ils sont dépourvus de cil ou de flagelle. Ils se déplacent grâce à des pseudopodes qui servent aussi à la capture des proies. La reproduction sexuée, dans ce groupe, n’est connue que chez les foraminifères.
3 Les Actinopodes.
Ils ont des pseudopodes à disposition rayonnante, soutenus par des filaments rigides (axopode). Leur forme est généralement sphérique.
B Les Apicomplexes / Sporozoaires.
Ils émettent des spores flagellées pendant leur cycle reproducteur. Ils n’ont pas d’appareil locomoteur. Ils sont généralement transmis par un vecteur (moustique). Exemple : Plasmodium falsiparum (paludisme).
C Les Ciliés (ou Ciliophores, ou Infusoires).
Ils présentent des cils à la surface de la cellule. Ils ont deux noyaux. Ils présentent divers modes de vie : libre (paramécie) ; fixé par un pédoncule ; symbiote ; parasites (peu nombreux).
D Les myxozoaires.
Ce sont des parasites de vertébrés, dont les poissons. En début de cycle, ils présentent une forme amiboïde qui évolue vers un plasmode plurinucléé : ils donneront une tumeur chez l’hôte. Le plasmode plurinucléé pourra aussi donner des spores complexes entourées d’une enveloppe de plusieurs cellules valvaires et donner finalement un germe pluricellulaire et plurinucléé.
IV Biologie des protozoaires.
A La locomotion.
Le mouvement orienté permet la recherche de nourriture, d’un abri, d’un nouvel habitat, d’un partenaire sexuel. On trouve trois types d’appareils locomoteurs.
1 Les pseudopodes.
Les pseudopodes sont des extensions cytoplasmiques temporaires pour la locomotion et la capture des proies. En général, des pseudopodes se rétractent pendant que d’autres se forment. Quatre formes de pseudopodes existent.
 Les lobopodes.
Les lobopodes.
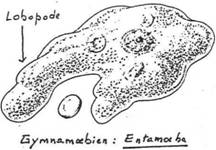
Ce sont des formes de digitation arrondie. Ils sont larges et courts, contiennent un endoplasme et un ectoplasme (périphérique). Les protozoaires présentant des lobopodes sont les amibes polypodiales (plusieurs pseudopodes) et les amibes monopodiales (un pseudopode).
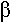 Les filopodes.
Les filopodes.
On trouve des filopodes chez les thécamoebiens (Difflugia). Ce sont des pseudopodes fins, parfois ramifiés mais qui pointent toujours à une extrémité de la cellule. Ils sont incapables de s’anastomoser.
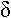 Les réticulopodes.
Les réticulopodes.
On les trouve chez les foraminifères. Ils sont fins, très ramifiés, se rejoignent pour constituer un réseau et même, les réticulopodes de plusieurs cellules peuvent se rejoindre et donner un réticulum multicellulaire (à filet pour piéger les proies). Exemple : Elphidium.
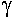 Les axopodes.
Les axopodes.
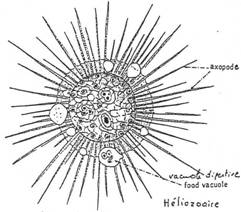
Ce sont des prolongements cytoplasmiques, à disposition rayonnante où chacun est soutenu par un filament axial (axonème) caractéristique des actinopodes.
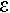 Formation du pseudopode.
Formation du pseudopode.
La formation du pseudopode résulte de l’existence de courants cytoplasmiques. Juste sous la membrane, on a une zone claire ; dans la zone interne, on trouve un endoplasme granuleux. Le plasmagel (ectoplasme) est un gel. Le plasmasol est interne et beaucoup plus fluide que le plasmagel.
La plasmagel applique une pression constante sur le plasmasol. Si en un point de la cellule, le plasmagel est insuffisant, le plasmasol va faire irruption et envahir le pseudopode en extension.
Quand le plasmasol atteint l’extrémité du tube, il fait demi-tour et se transforme en plasmagel. Très rapidement, à l’extrémité du plasmasol se forme une couche rétentrice qui arrête le mouvement.
è C’est le déplacement caractéristique des amibes nues. Leur déplacement est lent (2cm/H).
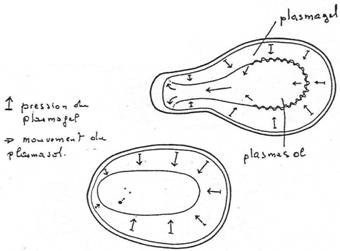
2 Cils et flagelles.
Les cils et flagelles sont permanents, en position fixe. Selon les cas, on les trouve sur toute la surface du corps ou localisés. Ils ne sont efficaces qu’en milieu fluide.
La locomotion par flagelles.
Ce type de locomotion caractérise les flagellés mais elle est aussi présente chez les spores et les microgamètes. Chez les flagellés, elle persiste chez les adultes. Les autres formes les perdent (cils et/ou flagelles) si elles s’enkystent.
La contraction du flagelle est une ondulation qui commence à la base et qui progresse vers l’extrémité.
En général, le déplacement se fait flagelle en avant, et la cellule semble tractée par son flagelle : c’est un mouvement de tractelle.
Si l’onde d’ondulation va de l’extrémité vers la base, la cellule est alors poussée et c’est un mouvement de pulselle que l’on observe (cas des spermatozoïdes).
En orientant le flagelle, il y a changement de direction.
Les flagelles permettent aussi un déplacement en latéral. L’onde de courbure peut être plane ou tridimensionnelle. Dans le denier cas, le flagelle fonctionne comme une hélice, mais provoque la rotation du corps autour de son axe.
La locomotion ciliaire.
Les cils se présentent sous forme de rangées, à la surface du corps.
Ils ont deux phases, effective et de recouvrement :
- Phase effective. Cette phase réalise la poussée. Le cil se raidie et se courbe à sa base pour donner un coup de fouet dans le plan de la rangée. Ce mouvement produit une impulsion. La direction de battement peut être inversée à tous moments.
- Phase de recouvrement (ou de récupération). Le cil retrouve sa position initiale. Il devient flasque et quitte le plan de mouvement pour se coucher sur le côté : pas de résistance à l’eau.
Quand il y a synchronie, tous les cils battent en même temps. Le plus souvent, le mouvement des cils est synchronisé avec les ondes locomotrices qui parcourent le corps : c’est la « métachronie ». è Les cils battent selon une séquence qui commence en un point et se propage sur le reste de la surface comme une vague.
L’onde locomotrice améliore l’efficacité du cil.
La surface est légèrement oblique par rapport au corps et l’onde parcourt un trajet en spirale autour de la cellule. è Les ciliaires se déplacent en s’enroulant autour d’un axe. Les paramécies se déplacent à 60cm/H.
Autres dispositifs.
- Les cils buccaux : ils entraînent les aliments vers la bouche.
- Les cils somatiques : ils servent à la locomotion.
Chez les ciliés hypotriches (Stylonychia), on trouve des groupes de 5 à 7 cils qui s’associent en petites touffes pour former des cirres. Dans ce cas, les cirres supportent le corps et permettent un déplacement sur substrat solide.
e Membranes ondulantes. Exemple chez un flagellé : Trypanosoma.
Chez Trypanosoma, le flagelle est replié le long du corps cellulaire, et, entre la membrane cellulaire et le flagelle, se forment des points d’accolement qui créent une véritable membrane ondulante (prolongement de la membrane cytoplasmique).
L’extrémité du flagelle est libre. Ce sont ses ondulations qui font bouger la membrane.
B Nutrition.
1 La prise de nourriture.
On trouve quelques saprophytes qui vont directement absorber les composés au travers de leur paroi : le système nutritionnel dégénère.
Les autres sont des holozoïques. Ils se nourrissent de nourriture solide (par prédation ou filtration).
La prédation.
Les protozoaires pratiquant la prédation sont très mobiles. Par exemple, une amibe eut attraper une paramécie : plusieurs lobopodes participent à la prédation. La proie est ingérable en n’importe quel point du corps. Les pseudopodes servent à capturer la proie.
Il y a ensuite libération de substance toxique pour immobiliser la proie, puis, mise en place d’une vacuole digestive.
Les paramécies ont une « bouche » (le cytostome), située au fond d’un entonnoir cilié (le cytopharynx).
L’entonnoir est garni de cils qui, en battant, dirigent les proies vers la bouche. Le cytopharynx a un grand nombre de trichocystes qui paralysent les proies. Ces dernières sont ensuite amenées dans la vacuole digestive.
Filtration ou « piégeage ».
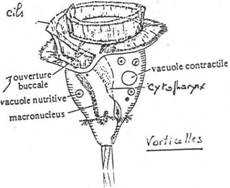
Ce mode de nutrition est souvent réservé aux organismes sessiles (fixés), par exemple, Vorticelles (péritriche) : sa couronne de cils, par des battements, crée des tourbillons qui amènent les particules dans la bouche. Au fond du cytopharynx, il y a formation d’une vésicule digestive. La nourriture entre dans la cellule par endocytose.
Chez les choanoflagellés coloniaux, le flagelle bat et entraîne l’eau vers la collerette.
Chez les organismes libres comme les Actinopodes flottants, il y a augmentation du rayon d’action par de nombreux axopodes rayonnant.
Chez les foraminifères, les réticulopodes ramifiés s’anastomosent et forment ainsi un piège à petits organismes.
2 La digestion.
La vacuole digestive est l’organite permettant la digestion intracellulaire. Celle-ci dérive du plasmalemme. Les enzymes digèrent les éléments phagocytés. Il ne reste plus que les déchets non assimilables (dans la vacuole).
Ce sont les courants d’eau plasmique (ou cyclose) qui favorisent le trajet des vacuoles digestives. Aucun trajet n’est défini, sauf chez les ciliés. Là, la vacuole a un trajet défini de sorte que la position de cette vacuole renseigne sur l’état de digestion des éléments ingérés.
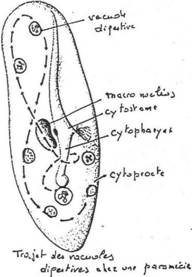
Pendant le trajet, il y a une série d’interventions d’enzymes, favorisées par les variations de pH.
Au début, le pH est neutre. Il devient ensuite acide (entre 3 et 4,5) puis remonte pour redevenir neutre à la fin du processus digestif.
3 Egestion.
La vacuole alimentaire entre en contact avec le plasmalemme et les déchets sont évacués par exocytose. Chez les ciliés, l’exocytose se fait toujours au même point : on parle alors « d’anus » ou de « cytoprocte ».
Chez les amibes, la technique est différente. Les vacuoles usées s’accumulent dans une « queue » (l’uroïde) qu’elles traînent puis qui est abandonnée.
C Respiration et circulation.
La majorité des protozoaires est aérobie (les anaérobies sont indépendants de l’O2). Les protozoaires aérobies n’ont pas d’organites spécialisés pour la respiration ; il y a diffusion d’O2 par la paroi cellulaire.
Les cils et flagelles, par leurs battements, favorisent les échanges.
La circulation est assurée par les courants cytoplasmiques ; le transport étant favorisé par les déformations de la cellule.
D Excrétion et osmorégulation.
Ces deux fonctions (excrétion et osmorégulation) sont liées.
Les paramécies, à leurs deux extrémités, possèdent des vacuoles pulsatiles. Elles battent en opposition de phase. Quand une est en diastole, l’autre est en systole. Elles évacuent, par une ouverture temporaire de la membrane, l’eau qui entre par osmose dans la cellule, à partir d’un milieu hypotonique (eau douce). Leur rôle est de maintenir la pression osmotique.
Si les paramécies sont dans un milieu isotonique, les pulsations s’arrêtent. Les vacuoles n’existent pas chez les protozoaires marins et parasites.
Les déchets solubles sont évacués avec l’eau rejetée par les vacuoles pulsatiles (en partie). La plus forte partie de l’excrétion est assurée par la membrane (à son travers), sans intervention d’organite.
Tout le tour de la cellule est en contact avec l’eau, ce qui facilite les passages.
Quand les protéines sont dégradées, les déchets sont de type azotés : les protozoaires sont dits ammoniotéliques. Les vacuoles digestives participent à l’exocytose.
E La reproduction.
Il existe deux types de reproduction chez les protozoaires : la multiplication asexuée et la reproduction sexuée.
1 La multiplication asexuée.
C’est le mode le plus répandu chez les protozoaires, surtout quand les conditions du milieu sont défavorables. Certains protozoaires ne pratiquent que cette méthode de multiplication.
La multiplication asexuée n’implique qu’un seul parent : tous les descendants seront donc identiques. Il existe alors un risque si le milieu est modifié (devient défavorable).
La fission binaire.
La cellule se divise en deux. C’est le type le plus courant (les protozoaires présentent deux à trois fissions binaires par jour). La fission peut être non orientée (comme chez les amibeslongitudinale chez les flagellés (Trypanosoma) ou transversale chez les paramécies (ciliés). [animaux sans forme précise]),
Il peut y avoir division du noyau sans division du cytoplasme : on obtient alors une forme transitoire ou résistante (Amoeba binucleata).
Si l’on trouve un grand nombre de noyaux, on parle de syncytium.
Le bourgeonnement (ou gemmiparité).
Il y a apparition à la surface cellulaire d’un bourgeon exogène, suivie d’une division nucléaire, capable de constituer un individu complet qui se détache de l’individu souche. èfission binaire inégale. C’est une
Un bourgeonnement dans le cytoplasme est appelé bourgeonnement endogène.
Les divisions multiples ou schizogonie.
C’est un phénomène courant chez les sporozoaires, qui existe chez les foraminifères. Il y a une division répétée du noyau puis des divisions du cytoplasme qui forment autant d’individus qu’il y a de noyaux.
Une masse de cytoplasme va être abandonnée, puis meurt.
2 La reproduction sexuée.
Il y a formation de cellules spécialisées (les gamètes) qui s’uniront en donnant un œuf (le zygote). Ce dernier est semblable morphologiquement aux parents mais génétiquement unique.
La reproduction sexuée assure une variabilité génétique de la population et donc, augmente la résistance de l’espèce aux conditions du milieu.
L’hétérogamie.
On trouve deux types de gamètes. C’est le mode de reproduction le plus répandu chez les protozoaires (reproduction amphimitique) car elle fait intervenir deux géniteurs. Il existe deux phases :
- La gamétogamie : c’est la formation des gamètes qui sont soit identiques morphologiquement (= isogamie, chez les foraminifères), soit différentes morphologiquement (= anisogamie, chez des sporozoaires). L’isogamie existe chez les protozoaires les plus primitifs.
- La gamontogamie : c’est l’appariement des deux gamontes (mâle et femelle) sans passer par un véritable gamète (les gamontes donnent les gamètes).
La conjugaison (uniquement chez les ciliés) ; exemple de la paramécie.
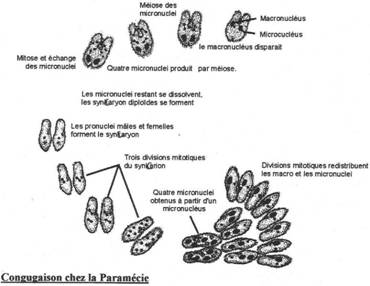
C’est une fécondation réciproque de deux individus accouplés qui échangent une partie de leur matériel nucléaire. Chaque conjuguant devient un vrai zygote. Il y a échange de noyaux haploïdes dont la fusion rétablie la diploïdie.
- Accolement au niveau du péristome à les macronucléus dégénèrent.
- Les micronucléus subissent une division équationnelle è deux cellules à noyau diploïde.
- Une division réductionnelle à deux cellules à quatre noyaux haploïdes è Les pronucléi.
- Trois pronucléi par cellule dégénèrent. Le quatrième subit une mitose et donne deux noyaux haploïdes par cellule : ce sont des noyaux de fécondation (un mâle et un femelle par cellule).
- Echange des noyaux mâles.
- Dans chaque cellule, les pronucléi fusionnent : c’est la fécondation.
- On obtient deux zygotes : le syncaryon.
Ensuite, il y a plusieurs divisions post-conjugaison qui rétablissent le complément nucléaire dont, la reformation du macronucléus dans chaque cellule fille.
L’autogamie.
L’autogamie est un mode de reproduction automictique : n’implique qu’un parent. Ce n’est pas une multiplication bien qu’un individu donne un individu. C’est un rajeunissement génétique de l’individu. On rencontre cette reproduction chez certains ciliés, foraminifères et héliozoaires (actinopodes).
Exemple du cycle d’Actinophys.
- Il rentre ses pseudopodes et s’enkyste.
- Il se divise et donne deux gamontes (un mâle et un femelle).
- Les gamontes donnent des gamètes.
- Les gamètes fusionnent en un seul individu.
è C’est un peu une modification de la conjugaison avec une simple redistribution des gènes.
3 Les cycles.
On distingue trois types de cycles schématiques biologiques des protozoaires.
Cycle haplobiontique.
La phase haploïde est longue. Seul le zygote est à l’état diploïde. La réduction chromosomique est immédiate.
Cycle haplodiplobiontique.
Il y a alternance des phases haploïde et diploïde qui sont équivalentes en durée.
La méiose a lieu à la fin de la vie de l’organisme diploïde è L’alternance de phase est une alternance de génération.
Exemple d’Elphydium crispum (foraminifère).
La génération diploïde présente un individu microsphérique alors que la génération haploïde montre un individu macrosphérique.
Cycle diplobiontique.
Les individus sont diploïdes. La méiose intervient lors de la gamétogenèse ou pendant la rencontre des individus chez les ciliés.
V Association avec d’autres organismes.
Les protozoaires épizoïques vivent fixés sur des plantes ou sur des animaux (sur la surface corporelle). Pour la nutrition, ils sont indépendants de l’hôte.
Les protozoaires endozoïques (qui vivent dans un autre organisme).
Ils sont totalement dépendants de leur hôte. On les trouve dans les organes creux, dans les tissus ou dans les cellules.
On distingue deux cas :
- Mutualisme et symbiose.
- Parasitisme.
A Mutualisme et symbiose.
Les protozoaires entretiennent un rapport à bénéfice réciproque avec leur hôte. On détaille deux cas :
Exemples d’association tripartite : protozoaire + bactéries + termite et protozoaire + bactéries + mammifères ruminants.
Dans tous les cas, les symbiotes sont localisés dans un segment du tube digestif qui devient une chambre de fermentation.
1 Association avec les termites.
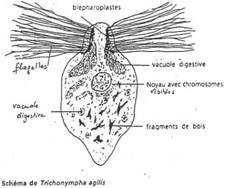
Les termites sont xylophages et ont par conséquent, un régime alimentaire pauvre en éléments azotés. Il se forme une chambre de fermentation dans un diverticule du proctodéum : la panse rectale.
Cette panse abrite une faune importante de Trichonympha et de bactéries. Trichonympha a un aspect piriforme. La partie apicale forme un rostre qui porte de nombreux flagelles. Ces protozoaires possèdent l’équipement enzymatique nécessaire qui permet la digestion de la cellulose (les termites en sont dépourvus).
è Les termites concassent le bois en fragments phagocytés par les Trichonympha, puis, digèrent la cellulose et rejettent de la lignine.
Au centre de ces cellules de protozoaires, on trouve un très grand nombre de bactéries qui entourent l’axostyle et les fragments de bois. Les bactéries dégradent les déchets puriques, ce qui restitue de l’azote aux termites.
Un termite sans Trichonympha meurt.
L’intérêt pour le protozoaire est qu’il est abrité par l’insecte et alimenté en aliments déjà mâchés. Cette symbiose est constante chez les termites et chez certaines blattes.
Le compartiment proctodéal du termite est soumis aux mues à perte des protozoaires. Le termite doit réensemencer son tube digestif en ingérant sa dépouille ou en quémandant des gouttelettes du contenu rectal d’un congénère.
2 Association avec les mammifères ruminants.
Les protozoaires sont capables de digérer la cellulose alors que les mammifères en sont incapables. La digestion de cette matière se fait donc dans un estomac compartimenté en quatre chambres digestives (panse, bonnet, feuillet, caillette). La panse est la chambre de fermentation. Chez le bœuf, elle peut atteindre 250 litres. On y trouve 1011 bactéries/mL et 106 ciliés/mL. Les bactéries digèrent aussi la lignine. Les protozoaires sont anaérobies.
Les ciliés phagocytent la lignine mais aussi les bactéries. Le bœuf digère de grandes quantités de ciliés et de bactéries.
On considère qu’un bœuf digère 1kg à 1,5kg de bactéries et de ciliés par jour.
Les ruminants sont un groupe prospère : leur réussite est sans doute liée à cette association symbiotique.
B Le parasitisme.
Un parasite est un organisme qui vit au dépend d’un autre être vivant. Pour le parasite, l’association est obligatoire.
1 Parasitisme chez les flagellés.
Exemple des Trypanosomides (reproduction sexuée inconnue).
Ils s’attaquent à l’Homme et au bétail. Certains se développent dans les parties antérieures du tube digestif d’un insecte piqueur. C’est le cas de Trypanosoma brucei gambience et de Trypanosoma brucei rodhésience. Ils provoquent la maladie du sommeil et sévissent soit en Rhodésie, soit en Gambie. Ils sont transmis par la mouche tsé-tsé ou Glossine. La transmission se fait par piqûre.
Le trypanosome peut vivre aussi chez des mammifères sans les inquiéter (porteurs sains) tels les antilopes et les porcs : ce sont des réservoirs à virus.
Exemple de Leishmania aethiopica.
Ce dernier est transmis par un petit moustique. Ses « réservoirs à virus » sont les chiens et les rongeurs. Quand l’Homme est piqué, le flagellé pullule au point d’inoculation. èbouton d’orient) qui laisse des traces indélébiles. On trouve ce parasite en Afrique du Nord et au Moyen-Orient. Altération cutanée (le
Exemple des Trychomonadine.
Ils ont une forme en amande et portent 4 à 5 flagelles répartis en une petite touffe apicalemembrane ondulante. et un flagelle formant une
Le cas de Trichomonas vaginalis. C’est un parasite du tractus uro-génital. Chez la femme il provoque un déséquilibre de ce tractus : inflammations ou vaginites.
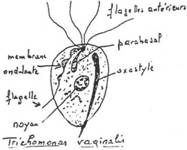
La transmission à l’homme se fait par voie sexuelle et ils s’installent dans l’urètre et la prostate où ils donnent des kystes. Ils peuvent survivrent plusieurs heures sur des linges souillés… Ce qui explique la « vaginite de la vierge ».
Chez l’homme, ils sont responsables de 15% des inflammations de l’urètre.
2 Parasitisme chez les rhizopodes.
Exemple d’un amibe : Entamoeba histolyca. (Amibe monopodiale).
On ingère un kyste avec de l’eau ou des légumes souillés par des excréments humains. Le dékystement se fait dans le gros intestin.
Il existe deux formes d’Entamoeba histolyca : une forme non pathogène (minuta) qui se nourrit de bactéries et d’amidon ; un forme pathogène qui va perforer les capillaires sanguins et sécréter des enzymes histolytiques qui provoquent des ulcères du colon. Il y a déclenchement d’une dysenterie amibienne caractérisée par des douleurs abdominales et des diarrhées sanguinolentes. Ces amibes peuvent ensuite gagner le foie, le cerveau, les poumons… en passant par le sang. On les trouve dans les zones tropicales.
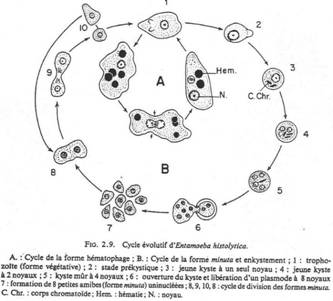
3 Parasitisme chez les sporozoaires.
Les sporozoaires sont tous parasites. Leur cycle est caractérisé par l’alternance d’une phase asexuée (schizogonie) et d’une phase sexuée (gamogonie). La schizogonie se déroule lors de l’infestation de l’hôte. La gamogonie permet la transmission d’un hôte à l’autre.
Le cycle débute avec le sporozoïte (qui contamine l’hôte définitif) qui donne un trophozoïte. Il y a ensuite multiplication des noyaux, divisions du cytoplasme, ce qui entraîne l’apparition d’autant de schiphozoïtes qu’il y a de noyaux. Les schiphozoïtes s’accroissent pour donner les gamontes (gamogonie).
Les gamontes femelles s’accroissent pour évoluer en macrogamètes. Les gamontes males subissent la gamogonie (division du noyau puis du cytoplasme) : au final, on a alors un grand nombre de gamètes males.
L’anisogamie est de règle.
La fusion des gamètes donne un zygote (diploïde) qui subit de suite la méiose.
La phase de sporogonie est caractérisée par des multiplications du noyau è Il y a libération d’un grand nombre de spores. Ces dernières donneront les sporozoïtes.
On observe deux principaux groupes : les Coccidies (de petite taille) et les Grégarines (de grande taille).
Exemple de Coccidie monoxème (maladie du « gros ventre » chez les lapins).
Le lapin s’infeste en mangeant de l’herbe souillée. Les spores ingérées arrivent dans le tube digestif et libèrent des sporozoïtes qui entrent dans les cellules intestinales. Les sporozoïtes se multiplient et vont infester d’autres cellules intestinales.
La fécondation a lieu dans la lumière du tube digestif. Les zygotes s’entourent d’une membrane épaisse : ce sont les spores. Le tout est rejeté avec les excréments.
Exemple de Coccidie dixène.
Les hôtes définitifs sont les félidés (chat, puma, jaguar…). Après ingestion, il y a infestation des cellules intestinales, puis, multiplication par schizogonie. Les cellules intestinales vont éclater et libérer des sporozoïtes qui parasiteront d’autres cellules. Les gamontes males et femelles vont fusionner pour former un zygote (oocyste) qui sera évacué avec la matière fécale.
Dans le milieu extérieur se produit la méiose. L’oocyste donne deux sporocystes contenant chacun 4 sporozoïtes. Le pouvoir infectieux va de 30 jours à sec, à 1 an en milieu humide.
Le passage à l’hôte intermédiaire est possible. Il y a enkystement dans les muscles.
Le passage à l’Homme se fait par des légumes souillés ou par le bœuf, le porc, le mouton… mais l’Homme n’est pas l’hôte définitif. S’il y a prolifération des kystes, elle se fait au niveau des viscères, des poumons, des muscles et de l’encéphale.
La transmission congénitale est possible par voie placentaire. Si la contamination se fait durant le premier trimestre, il y a un risque d’avortement ou de malformation. Après ce premier trimestre, la contamination sera détectable par des signes d’infection généralisée à la naissance.
Coccidie hétéroxène (paludisme).
Le paludisme tue chaque année 2,5 millions de personnes.
La schizogonie a lieu chez l’Homme. La gamogonie et la sporogonie se font chez le moustique (anophèle). Les symptômes sont des accès de fièvre toutes les 48 heures, dues à l’éclatement des globules rouges et à la libération de protéines et du parasite dans le plasma.
Les Grégarines.
Les Grégarines parasitent les tubes digestifs d’invertébrés (annélides et insectes). Le trophozoïte est mobile (pour un déplacement dans l’intestin).
Exemple de Stylocephalus langicollis.
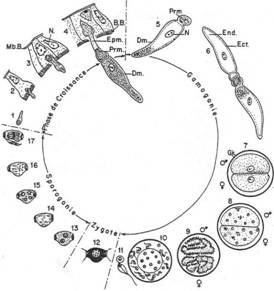
C’est un parasite d’un coléoptère du genre blaps.
L’infestation se fait par consommation de spores abandonnées par un individu atteint.
Dans les phases 1 à 3, le parasite s’incruste dans le cytoplasme des cellules de la paroi de l’intestin. Rapidement, le trophozoïte se différencie en trois parties :
- Une partie étroite, terminée en suçoir enfoncée dans les cellules intestinales : l’épimérite.
- La partie moyenne : le protomérite.
- La partie externe : le deutomérite. Il renferme la plus grande partie du cytoplasme et le noyau.
Le départ se fait par rupture de l’épimérite qui reste dans la cellule hôte. Ensuite, il y a accolement, deux à deux, des trophozoïtes (= syzygie) au niveau de protomérites è mucus qui entoure les deux cellules (une cellule mâle et une cellule femelle è petit kyste appelé gamétocyste, évacué avec les fécès).
Dans le milieu externe, les noyaux se multiplient et se disposent à la périphérie du cytoplasme
Phase 9 : perlage des gamètes.
Après la fécondation, on obtient un grand nombre d’œufs. Un zygote donne une spore qui subit trois divisions successives (la première étant réductionnelle).
4 Parasitisme chez les Myxozoaires.
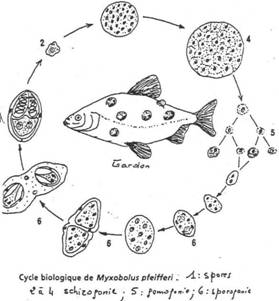
Exemple de Myxobolus pfeifferi.
En 2, on a un germe amiboïde : le sporoplasme.
En 1, on a le stade final : la spore contenant le germe.
Le sporoplasme est binucléé (2 noyaux). La structure de la spore est pluricellulaire è on a une coque périphérique à plusieurs cellules. Elle renferme le sporoplasme et un filament, dont le déroulement favorise la libération de sporoplasme.
Le poisson s’infeste en ingérant les spores. Elles s’ouvrent dans l’intestin et libèrent le sporoplasme qui va se localiser dans les muscles où il subira de nombreuses schizogonies. Le tissu hôte donne des tumeurs qui abritent un grand nombre de kystes à structure en plasmobe.
On suppose que dans le plasmobe, il y aurait réduction chromatique et gamétogenèse. Ce qui est sûr, c’est qu’il y a évacuation d’une spore avec les excréments.
VI Les formes coloniales.
Des colonies existent chez les choanoflagellés : elle varient de quelques individus à plusieurs milliers. Il peut même y avoir une différentiation des individus (végétatifs/reproductifs).
Par ces adaptations, les protozoaires coloniaux se rapprochent de l’organisation des pluricellulaires.
Chapitre 2 :
Les Diploblastiques.
Les diploblastiques sont, de façon générale, des métazoaires : une association de cellules qui se partagent le travail. Il existe de nombreux stades chez les métazoaires.
Il existe un groupe, les parazoaires, situés entre les protozoaires et les métazoaires : les spongiaires. Ces spongiaires sont une association cellulaire pouvant se défaire. La différenciation n’apparaît pas d’emblée. Chez les diploblastiques, il existe des cellules totipotentes.
Lorsqu’une partie du génome est exprimée et que l’autre partie est inhibée, on a une cellule différenciée.
Les cellules différenciées s’associent pour former des couches monostratifiées : les feuillets. Quand les cellules forment les deux feuillets fondamentaux, on parle alors d’organismes diploblastiques. Le feuillet interne est appelé endoderme et le feuillet externe est appelé ectoderme.
Les mêmes phénomènes sont observés au cours de l’embryogenèse. Le début d’un animal est un stade unicellulaire (l’œuf) qui passe ensuite par le stade diploblastique (gastrula). Chez les diploblastiques, le développement s’arrête à ce stade.
Les trois embranchements des diploblastiques sont : les spongiaires, les cnidaires (polypes et méduses) et les cténaires.
I Les spongiaires.
Les spongiaires sont des organismes essentiellement marins. On trouve autour de 5000 espèces. Ce sont des organismes sessiles (vivant fixés). Leurs seuls mouvements sont des contractions locales du corps et des mouvements d’ouverture et de fermeture des pores.
A Organisation.
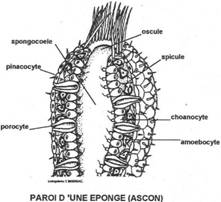
Ce sont donc des parazoaires. Les différentes catégories cellulaires n’ont pas de cohésion définitive. Dans la mésoglée, on trouve de nombreuses cellules totipotentes.
Le spongiaire type est le type olynthus : c’est une amphore fixée par sa base. L’apex porte un orifice exhalant unique (l’oscule) et la paroi est percée de nombreux pores inhalant.
Suivant la complexité de la paroi, on distingue différents stades (pas de groupe systématique).
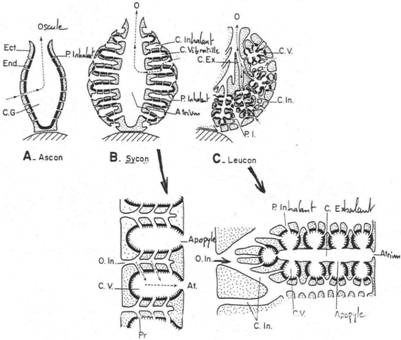
1 Le stade Ascon.
Le stade ascon est le stade le plus juvénile de la plupart des éponges. La structure de la paroi est identique en tous points du corps. Il n’y ni organe ni appareil.
La mésoglée forme une gelée où circulent différentes catégories cellulaires, totalement indépendantes.
L’ectoderme est formé d’un épithélium de revêtement : les pinacocytes. Ce sont des cellules aplaties, jointives, recouvertes d’une fine cuticule. Entre les pinacocytes s’ouvrent les pores inhalants, formés par des porocytes (pour l’entrée d’eau).
Le feuillet interne tapisse toute la cavité gastrale (ou atrium ou spongocœle) et est composé de choanocytes. Ce sont des cellules de grande taille, pourvues d’un long flagelle et l’apex forme une collerette de nombreuses microvillosités. Les choanocytes ressemblent aux choanoflagellés. Ils créent un courant d’eau dans l’atrium, ce qui permet la capture de particules alimentaires dans la collerette, mais aussi, la circulation de l’O2.
Dans la mésoglée, on trouve :
- Des scléroblastes : ils sécrètent les spicules. Ils peuvent se grouper par groupe de 2 ou 3 (et donner ainsi des spicules de 2 ou 3 axes).
- Des cellules amiboïdes à rôle phagocytaire.
- Des collancytes qui sécrètent la mésoglée.
- Des archéocytes : ce sont de véritables cellules totipotentes.
- Des myoblastes (en petit nombre) : ce sont des cellules capables de contraction.
- Des cellules nerveuses, diffuses.
2 Le stade Sycon.
Les choanocytes se localisent dans des diverticules tubulaires qui débouchent dans l’atrium par des orifices : les apopyles. è Il y a formation de canaux pour piéger l’eau chargée de particules alimentaires.
3 Le stade Leucon.
Chaque diverticule se divise en diverticules secondaires appelés corbeilles vibratiles. Les choanocytes sont dans ces corbeilles. Elles débouchent dans des canaux exhalants, en relation avec l’oscule.
B Classification.
On reconnaît trois classes, selon la nature des spicules :
- Les éponges calcaires : les spicules sont composés de calcite. Elles peuvent être simples ou composées. Il existe deux formes :
· Homocœle : les choanocytes tapissent tout l’atrium.
· Hétérocœle : il y a formation de corbeilles vibratiles ou de diverticules tubulaires.
- Les Hexactinellides (ou triaxonides ; genre Euplectella). Les spicules sont composées de silice hydratée et donnent une architecture cohérente. Elles possèdent trois axes et parfois, peuvent atteindre une longueur de 60 centimètres.
- Les Démosponges : le squelette est formé de spongine pouvant être associée à quelques spicules siliceux.
C La reproduction et le développement.
Les éponges sont gonochoriques ou hermaphrodites ; il n’y a jamais de gonades.
1 La gamétogenèse.
Les gamètes mâles.
Ces gamètes sont des archéocytes diploïdes situés dans la mésoglée. Ils se divisent en deux cellules diploïdes dont l’une entoure l’autre. Ensuite, il y a méiose et se forme 4 cellules haploïdes qui seront les spermatozoïdes.
Les gamètes femelles.
Ces gamètes sont dans la mésoglée. Les archéocytes se différencient en cellules arrondies (oogonies) qui passent dans les corbeilles vibratiles. C’est là qu’à lieu la méiose. Les cellules regagnent la mésoglée et connaissent une augmentation du volume cytoplasmique et sont alors des ovocytes.
2 La fécondation.
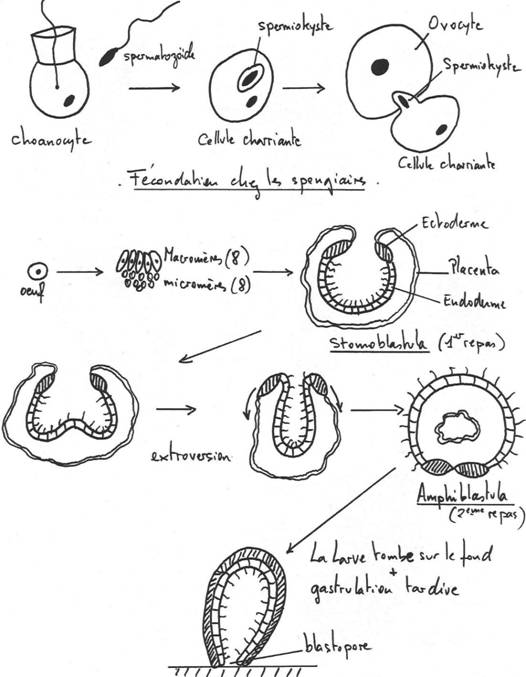
Un spermatozoïde pénètre dans un choanocyte et perd son flagelle. Le choanocyte perd collerette et flagelle. Le spermatozoïde devient alors un spermiokyste alors que le choanocyte devient une cellule charriante.
Cette dernière va aller au contact de l’ovocyte pour y injecter le spermiokyste.
C’est une fécondation indirecte.
3 Le développement embryonnaire et larvaire.
Le développement est particulier. Les éponges, vivipares, incubent l’embryon dans la mésoglée. Après la fécondation, l’œuf subit 4 divisions qui l’amènent au stade 16 blastomères (8 macromères et 8 micromères). Les macromères donneront les cellules supérieures à potentialité ectodermique alors que les micromères ont une potentialité endodermique.
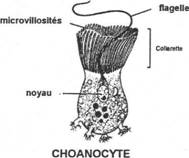
Les micromères se divisant plus vite que les macromères, on atteint un stade intermédiaire (blastula) : sphère creuse. Les cellules endodermiques de la blastula vont acquérir des flagelles alors que les cellules ectodermiques s’écartent pour ménager un orifice (différent du blastopore). è C’est le stade stomoblastula. Le premier repas se fait par consommation des choanocytes de la mère.
La stomoblastula subit une inversion des feuillets par extroversion : inversion des surfaces par l’ouverture ménagée dans l’ectoderme.
è On obtient une larve nageuse typique ou amphiblastula.
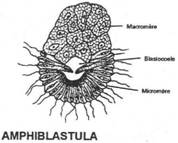
Lors du mouvement de migration des feuillets, le placenta s’est retrouvé dans la larve : il se fait digérer ; c’est le second repas embryonnaire.
La larve va tomber sur le fond et il se passe alors la véritable gastrulation. La larve se fixe par le blastopore, l’endoderme perd sa ciliature alors que se différencient les choanocytes.
Le blastopore se ferme et l’oscule se perce à l’apex. L’ectoderme voit se différencier des sporocytes.
4 Notion d’individu.
C’est une notion floue car les cellules sont très malléables. Si l’on fragmente une éponge, on peut faire un bouturage. Il peut se former des amas d’archéocytes affleurant en surface. Ils se détachent (en devenant des propagules) et donnent un nouvel individu. Les propagules peuvent être différenciés.
Les propagules s’entourent de deux couches cellulaires qui édifient des amphidisques. Ces derniers vont attendre le retour de conditions favorables pour redonner des propagules puis de nouveaux individus.
à Il n’y a pas d’individu au sens strict, ni de colonie.
è Ce sont des individus particuliers (parazoaires).
II Les cnidaires.
Les cnidaires sont à 99% marins, 1% d’eau douce (cnidaire = radiata). Il existe une symétrie radiaire, plusieurs tentacules entourent la bouche. Ces animaux ont une cavité gastro-vasculaire interne avec un seul orifice (la bouche).
Deux formes peuvent alterner pour la même espèce :
- La forme méduse : elle est libre, pélagique et constitue (généralement) la forme sexuée.
- La forme polype : elle est benthique, fixée. C’est une forme asexuée (généralement).
Les polypes peuvent être solitaires ou coloniaux et acquérir alors, un polymorphisme des individus : c’est une variation morphologique intraspécifique liée à la spécialisation des individus.
On note la présence de cellules urticantes : les cnidocystes (ou cnidoblastes, ou nématoblastes, ou nématocystes). Ces cellules contiennent un filament urticant baignant dans une toxine (l’actino-congestine).
Les cnidoblastes ont un cnidocil qui, lorsqu’il est excité, entraîne la dévagination du fil urticant qui peut tuer la proie ou créer des irritations.
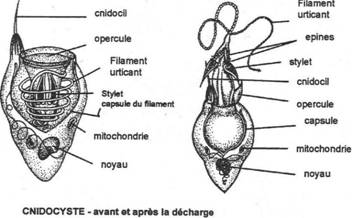
La paroi possède une structure diploblastique. Ectoderme et endoderme sont séparés par la mésoglée.
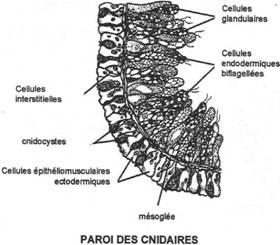
Au niveau de l’ectoderme, il y a un pavage de cellules épithélio-musculaires, des cnidoblastes et quelques cellules interstitielles totipotentes.
L’endoderme est constitué de cellules biflagellées à rôle phagocytaire et de cellules glandulaires sécrétant les enzymes digestifs (la digestion se déroule en deux temps). Il n’y a pas fabrication d’organe au sens strict.
La reproduction sexuée produit un œuf, puis, à l’éclosion, on obtient une larve ciliée nageuse (planula) à polarisation antéro-postérieure (avec donc, une symétrie bilatérale). La symétrie radiaire est la conséquence de la fixation et de la métamorphose de la planula.
Il existe trois classes de cnidaires : les anthozoaires, les hydrozoaires et les scyphozoaires.
A Les Anthozoaires.
Seule la forme polype persiste : l’organisation du polype se complique donc. Il existe une symétrie bilatérale déterminée par une invagination ectodermique qui forme le pharynx (ou stomodeum). Le pharynx est aplati latéralement.
La cavité gastrale voit l’ectoderme former des replis qui déterminent des cloisons. Dans la région antérieure, ces cloisons se soudent au pharynx : on a alors des loges. Il y a autant de tentacules que de loges.
On distingue deux groupes :
- Les octocoralliaires : ils ont 8 tentacules, 8 loges et 8 cloisons.
- Les hexacoralliaires : ils ont n´6 tentacules, n´6 tentacules et n´6 cloisons.
1 Les octocoralliaires.
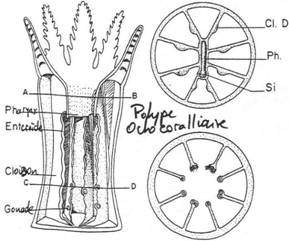
Chaque tentacule porte deux rangées de pinnules et est muni de cnidoblastes.
Le pharynx a une section plus ou moins ovale et est muni, à une extrémité, d’une gouttière ciliée : le siphonoglyphe. Le battement des cils permet la progression des particules alimentaires vers la cavité gastrale. Cette dernière possède 8 cloisons à disposition radiaire.
Les octocoralliaires sont tous coloniaux et les individus de la colonie sont tous semblables. Ils sont tous issus d’un même polype souche, lui-même issu de la fixation de la planula. La formation de la colonie est réalisée par bourgeonnement.
Les différents polypes sont unis par des évaginations tubulaires (les stolons). Ce sont sur ces stolons que bourgeonnent les nouveaux individus. à Les colonies seraient peu importantes s’il n’y avait pas de squelette. è Le système de construction du squelette permet de différencier les différentes formes d’octocoralliaires.
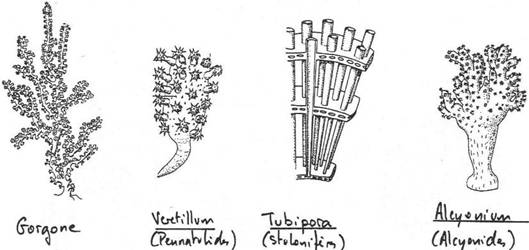
Les Alcyonides.
Dans la mésoglée, les scléroblastes forment des spicules creux qui s’enchevêtrent pour former une structure solide. Le squelette est peu dur et la colonie a un aspect digité. Les polypes sont reliés par des tubes endodermiques.
Les Stolonifères (Tubipora = « orgue de mer »).
Les stolons ont une disposition très régulière et le squelette est constitué de spicules dans la mésoglée. Ces spicules ont une disposition très cohérente.
Les Gorgonides.
La colonie a un aspect arborescent ou en éventail. Elle est soutenue par un squelette externe corné pouvant être imprégné de sels calcaires.
Les Coralides (ex : le corail).
Les tubes endodermiques se ramifient dans la colonie et le squelette est calcaire, très dur et externe. Actuellement, les Gorgonides et les Coralides sont parfois regroupés ensemble, on parle uniquement de Coralides.
Les Pennatulidés.
La colonie se développe sur un axe (ou rachis), enfoncé dans le sable par sa base. Il se couvre de polypes. Le squelette est formé de spicules mésogléens.
2 Les Hexacoralliaires.
Les hexacoralliaires ont un système radiaire d’ordre 6. Leurs tentacules sont sans pinnule et les polypes peuvent être solitaires ou coloniaux. Lorsqu’ils sont coloniaux, ils forment des supports immenses : les récifs.
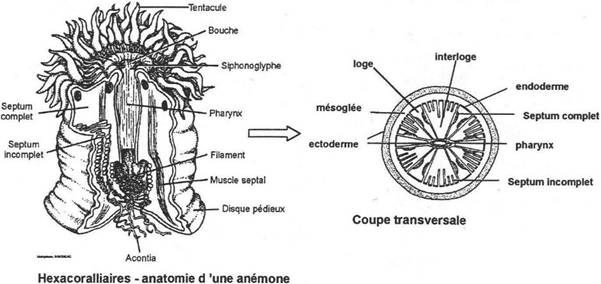
Les hexacoralliaires n’ont jamais de scléroblaste mésogléen ; par contre, ils possèdent des calcoblastes situés à la base de l’ectoderme (position basale).
Il existe deux types morphologique : les Actiniaires (mous) et les Sclératinides (formant les récifs).
Les Actiniaires.
Ces sont les anémones de mer, avec un polype solitaire et géant. Les diverses loges communiquent entre elles par des ostioles. Les Actiniaires ont aussi de longs filaments branchés sur l’endoderme (les aconties). Ces aconties sortent des orifices de la paroi (les cinclides) et sont garnies de cnidoblastes.
Les gonades apparaissent périodiquement.
La planula tombe sur le fond et donne un petit polype qui grossit par développement de la structure des cloisons.
Le jeune polype possède 12 cloisons macrentériques (principales) qui déterminent des loges puis des interloges. Au niveau des interloges se forment des cloisons micrentériques (ce sont des loges incomplètes).
Avec le temps, le nombre de tentacules s’accroît.
Les modes de vie sont très variés chez les Actiniaires :
- Anémones pivotantes : fouisseuses dans le sable.
- Anémones flottantes : piègent l’air dans un repli de leur base et flottent à l’envers.
- Anémones benthiques fixées.
- Anémones en association avec des pagures : elles sont fixées sur la coquille voir même sur les pinces. Elles bénéficient des restes des proies du pagure mais elles lui servent pour tuer les proies.
En dehors de la reproduction sexuée, il y a possibilité de scissiparité par découpage longitudinal ou latitudinal.
Les Madréporaires.
Les madréporaires sont dans les eaux riches en calcaire, bien aérées avec une température supérieure à 20°C.
A la base du polype apparaissent des lignées radiaires de calcoblastes : ils forment dans un premier temps une base homogène (plaque). Dans un second temps, ils sécrètent des lignées radiaires qui donnent des axes rayonnés.
Si les polypes sont accolés, il se forme un ensemble massif (genre Astrea). S’ils s’alignent et fusionnent partiellement, c’est le genre Meandrina.
B Les Scyphozoaires.
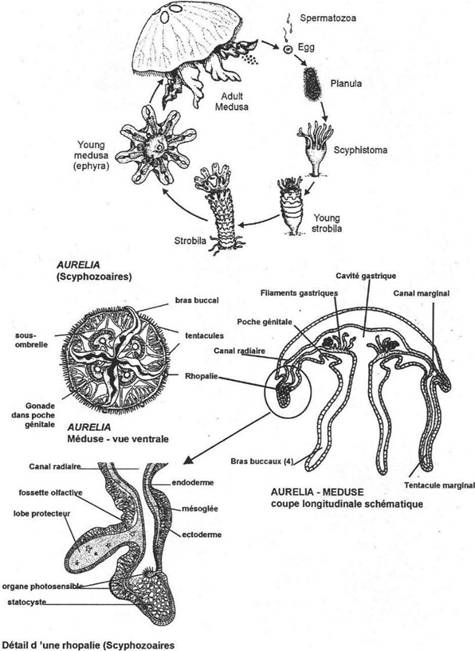
Dans cette classe, il y a simplification du polype : il devient une phase larvaire. Il est suivi de l’apparition de la méduse qui formera les gamètes. Le cycle typique est celui d’Aurélia.
La planula est typique, à deux feuillets. Elle tombe sur le fond, se fixe par la région antérieure pour se transformer en un petit polype scyphostome (jeune polype). Ce polype possède un orifice buccal. Il acquière ensuite 4 tentacules puis 8. Il mesure quelques millimètres.
La bouche se situe à l’extrémité d’une petite trompe : le probocis. La cavité gastrale est cloisonnée et possède quatre loges. Ce polype subit la multiplication asexuée par strobilisation : il subit une série de constrictions transversales pour prendre l’aspect d’assiettes empilées. Chacune de ces « assiettes » se détache et se retourne pour donner une jeune méduse (méduse éphyra). Sa croissance donnera la méduse adulte qui possède une série de tentacules courts, périphériques (formant des franges) et quatre bras buccaux. On trouve quatre 4 massifs gonadiques et des zones sensorielles périphériques : les rhopalies. Ces dernières possèdent des organes photosensibles (ocelles), le statocyste qui détecte la gravité (pour l’équilibration). La rhopalie est protégée par un lobe qui forme une sorte de bouclier. La cavité sous-ombrellaire est complètement ouverte (pas de vélum) : méduses acraspèdes ou méduses acraspédotes. Cette cavité est capable de contraction mais le déplacement sur de longues distances reste passif.
C Les Hydrozoaires.
On distingue divers groupes comme les Hydrides et les Leptolides. Les individus de ces deux groupes sont dépourvus de pharynx et la bouche s’ouvre directement à la surface du corps dans la cavité gastrale. Dans le cycle vital, il y a alternance des deux phases : polypes et méduses, sauf chez les Hydrides où la méduse disparaît et le polype est alors capable de reproduction sexuée et asexuée.
1 Les Hydrides.
On distingue des polypes mâles et femelles qui donnent des gamètes qui, par fécondation, donnent un œuf qui va tomber et donner un jeune polype (capable de multiplication asexuée par bourgeonnement) sans passage par le stade planula.
2 Les Leptolides.
Chez les Leptolides se développent des structures coloniales ; le stolon de bourgeonnement est appelé hydrorhize ; la base du polype est l’hydrocaule (» la tige) ; chaque polype est un hydrante. L’ensemble de ces formations charnues compose le cœnosarc. Ce dernier est protégé par une enveloppe d’origine ectodermique (le périsarc). Autour de chaque polype se forme une enveloppe protectrice (l’hydrothèque).
Dans le groupe des Calyptoblastides, l’hydrothèque enveloppe tout le polype alors que dans le groupe des Gymnoblastides, l’hydrothèque n’enveloppe que la base du polype.
Le polymorphisme permet de distinguer :
- Des gastrozoïtes (polypes nutritifs) qui ont une bouche et une couronne de tentacules.
- Des dactylozoïtes (polype défensif) riches en cnidoblastes, sans bouche ni tentacule.
- Des gonozoïtes qui réalisent le bourgeonnement des méduses. L’axe de bourgeonnement est le blastostyle.
Les méduses portent les gonades qui donneront les gamètes qui, par fécondation, donnent un œuf qui donne une planula qui va tomber sur le fond et former une colonie à partir du premier polype.
3 La formation des méduses.
- A Il y a formation de protubérances à la surface des blastostyles (= bourgeons médusaires), délimitées par les deux feuillets.
- B Il y a formation d’un nodule médusaire qui s’isole de l’ectoderme périphérique. Le nodule va se creuser d’une cavité.
- C+C’ Il comprend l’ébauche de la cavité sous-ombrellaire et celle des tentacules.
- D La lame didermique forme le vélum et une saillie axiale de la cavité va former le manubrium qui se perce de la bouche.
- E Le pédoncule de fixation s’étire et se rompt : c’est la libération de la méduse. Quand elle est détachée, elle réalisera la dissémination des gamètes. Toutes les méduses issues d’une même colonie ont le même sexe.
La multiplication asexuée peut atteindre la méduse :
- par scissiparité dans l’axe du manubrium ;
- par des bourgeonnements au niveau du manubrium.
4 Les Siphonophores.
Les Siphonophores sont des animaux pélagiques où la colonie est constituée d’une association de plusieurs unités (à cormidies). La cormidie est formée d’une association de polype polymorphiques.
Cette colonie se développe à partir de la planula. Cette dernière ne se fixe pas : à son pôle antérieur se forme un nodule médusaire qui va constituer le pneumatophore (cavité sécrétant un gaz riche en azote, oxygène et argon) qui a un rôle de flotteur.
La région postérieure s’allonge et forme un filament pêcheur et le tout premier gastrozoïte : c’est la larve siphonula.
Entre le premier gastrozoïte et le pneumatophore, on trouve une zone de croissance (le nectosome) qui permet la formation des cormidies.
La région antérieure du nectosome forme d’abord plusieurs bourgeons qui se différencient en cloches natatoires. Dans la région inférieure, les cormidies successives apparaissent.
Une cormidie comprend des gastrozoïtes, des dactylozoïtes (rôle défenseur, avec un palpe allongé), un filament pêcheur riche en cnidoblastes, des gonozoïtes mais aussi des individus médusoïdes, capables de se différencier en n’importe quel type polypier.
L’ensemble de la colonie est protégé par un bouclier. è Exemple : Halistemma.
Il existe d’autres types morphologiques :
- Les Anectes (Chryslia) : le pneumatophore forme un flotteur très développé muni d’une carène. Les cormidies sont directement fixées au flotteur et ne sont pas protégées par un bouclier.
- Les Chondrophorides (Vellela) : Ils utilisent le polymorphisme comme des organes : on trouve un seul gastrozoïte central entouré par une zone concentrique de gonozoïtes, eux-mêmes entourés de dactylozoïtes. Le flotteur est divisé en cloisons remplies d’air. La zone de mésoglée est riche en réserves (à le « foie »). La partie supérieure forme une voile qui permet de dériver avec le vent.
III Les cténaires (= les Cténophores).
Les cténaires sont marins et n’ont aucun lien phylétique avec les cnidaires. Ils sont pélagiques et carnivores.
On trouve un type morphologique par espèce. Ils ont une double symétrie bilatérale : un plan tentaculaire et un plan pharyngien (le deux sont perpendiculaires).
L’organisme type est le cydippe (son diamètre est compris entre 1,5 et 15 millimètres). Le pôle oral porte la bouche. Le pôle aboral porte un statocyste (organe d’équilibration).
Ces cténaires ont également deux tentacules (qu peuvent atteindre 15 centimètres). Ces derniers peuvent se rétracter dans des poches tentaculaires. A la surface du corps, on distingue huit rangées de palettes vibratiles (peignes ou cténidies). Leurs battements permettent le déplacement de l’animal.
Les tentacules portent des cellules particulières : les colloblastes, qui sont constitués d’une masse de granules adhésifs et d’un prolongement cytoplasmique qui forme le filament spiralé. Le noyau, rectiligne, forme un filament droit. Quand cette cellule est excitée, elle sort de l’ectoderme où elle reste accrochée par le filament spiralé et se sert des granules adhésifs pour attraper la proie.
Il existe deux autres formes anatomiques :
- Les Tentaculés (Cestus) : ce sont des cténaires dilatés latéralement dans le plan tentaculaire.
- Les Atentaculés (Beroe) : ils sont en forme de cloche avec une dilatation du pharynx
Les Métazoaires Triploblastiques
Acœlomates et Pseudo-cœlomates.
Chez les métazoaires triploblastiques, il y a apparition du mésoderme (troisième feuillet embryonnaire) qui dérive de l’endoderme pendant le développement embryonnaire. L’ectoderme et l’endoderme ont des fonctions bien définies ; le mésoderme permet la différenciation d’organes internes (appareil excréteur, organes génitaux) dont la musculature qui permettra des mouvements orientés (locomotion). Il apparaît le déplacement dans une direction donnée : les animaux ont maintenant un avant et un arrière.
· Apparition de la symétrie bilatérale.
La symétrie bilatérale remplace la symétrie radiaire des cnidaires ; l’animal a donc une région antérieure, une postérieure et une orientation dorso-ventrale. Cette symétrie existe à l’état embryonnaire mais peut disparaître chez l’adulte comme chez les oursinspentaradiés) ou chez les gastéropodes. (
Cette symétrie bilatérale est un des principaux facteurs qui vont infléchir le cours de l’évolution animale.
Le déplacement se fait tête vers l’avant.
· La céphalisation.
La céphalisation est un développement des appareils sensoriels et du système nerveux àorganes sensoriels se regroupent dans la région antérieure, au voisinage d’un système nerveux principal (encéphale) è C’est le développement de la tête s’il y a un « cerveau », des organes sensoriels et d’une bouche. elle favorise la coordination des mouvements. Les
Chez les triploblastiques, il existe deux groupes : les acœlomates et les cœlomates. Ces groupes sont différenciés par la structure du mésoderme :
- Si le mésoderme donne un tissu épais, on se trouve chez les acœlomates.
- Si le mésoderme donne un tissu creux favorisant l’apparition de vésicules cœlomiques, on se trouve chez les cœlomates.
I Les Acœlomates.
Le mésoderme vient remplir les espaces entre les organes : c’est un parenchyme.
A Les Plathelminthes.
Les plathelminthes sont les triploblastiques les plus primitifs. Ils présentent un aplatissement dorso-ventral avec, évidemment, l’existence des faces dorsale et ventrale. Le corps est organisé en deux territoires différents :
- La région céphalique, antérieure (fonctions sensorielles).
- La région postérieure, caudale.
Il n’y a pas d’appareil circulatoire vrai : donc, pas de sang. Il existe toutefois un liquide présent dans l’espace du parenchyme (l’hémolymphe).
L’appareil respiratoire est absent : les échanges gazeux s’effectuent à travers le tégument de l’animal, ou bien, comme chez les parasites, il n’y aura pas d’échange gazeux.
L’appareil digestif est soit incomplet, soit absent. S’il est incomplet, c’est qu’il lui manque l’anus à « appareil en cul de sac ». S’il est absent, les échanges seront possibles par osmose (pour les parasites).
Les plathelminthes comprennent environ 50 000 espèces et peuvent être libres ou parasites. Il existe six classes mais nous n’en verrons que 4 ici : Turbellariés (planaires), Trématodes (douves), Cestodes (ténia) et Monogènes (polystomiens).
1 Les Turbellariés.
Les turbellariés sont essentiellement aquatiques (eau douce / marin) avec quelques formes terrestres (dans les forêts tropicales humides). Leur taille est souvent inférieure à 2 centimètres et peuvent exceptionnellement atteindre 60 millimètres.
Ils sont libres et non segmentés. Ils présentent une région antérieure avec deux yeux (ou plus) et des auricules. Ils ont une région spécialisée au moment de la reproduction.
Le tégument.
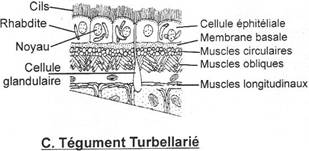
L’épithélium des turbellariés est cilié, épidermique. Le battement des cils sur l’épithélium permet la locomotion. Sous cet épithélium, on trouve une membrane basale puis différentes couches musculaires (circulaire, oblique, longitudinale).
Entre les couches musculaires, on distingue des cellules glandulaires qui peuvent émettre des digitations vers l’épithélium. Au niveau de la membrane basale, on a localement des cellules sensorielles.
Les cellules épithéliales ont un petit corpuscule (rhabdite) à rôle encore inconnu. Elles produisent un mucus à l’extérieur de l’animal pour enrober les corps étrangers.
Sous les couches musculaires, on a deux types cellulaires :
- Des cellules amiboïdes qui permettent les échanges respiratoires et nutritionnels.
- Des cellules étoilées qui baignent dans l’hémolymphe.
Système nerveux et organes sensoriels.
Les turbellariés sont les premiers animaux à posséder un système nerveux central bien défini. Ce système nerveux central comprend deux ganglions cérébroïdes réunis par une commissure du même nom. De ces ganglions partent les cordons nerveux latérauxnombreuses commissures transversales. (longitudinaux) et de
Des nerfs sensitifs partent aussi en direction des yeux et des auricules.
Au niveau des yeux est présent un tégument transparent qui permet l’entrée de la lumière. L’œil est composé d’une cupule tapissée de cellules pigmentées. Cette cupule concentre la lumière vers les cellules rétiniennes. è L’œil a une disposition inversée.
Les auricules ont un rôle tactile et parfois de préhension.
Il existe cinq récepteurs :
- Les tangorécepteurs : ils ont un rôle tactile lors des contacts avec des surfaces solides.
- Les rhéorécepteurs : ce sont des cellules sensorielles appréciant les courants.
- Les chémorécepteurs : ils détectent les composés chimiques.
- Les statorécepteurs : ils ont un rôle dans l’équilibration.
- Les photorécepteurs : ils sont sensibles à la lumière.
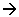 On ne trouve jamais les cinq types d’organes sensoriels à la fois dans le même animal.
On ne trouve jamais les cinq types d’organes sensoriels à la fois dans le même animal.
Exemple de statorécepteurs : les statocystes. Ils ont la forme d’une vésicule avec un statolithe et autour, une cellule spécialisée : un lithocyste.
L’appareil digestif.
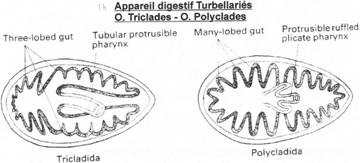
Un orifice (bouche) s’ouvre dans le 1/3 postérieur de l’animal. Cette bouche est suivie d’un pharynx d’origine ectodermique ; il est musculeux et peut être dévaginable. Après le pharynx, on a l’intestin, très volumineux, pouvant être ramifié.
Il existe trois types de pharynx :
- Le pharynx simple : il est formé par une invagination de l’ectoderme (à épithélium cilié) avec une fine paroi musculaire.
- Le pharynx bulbeux : il est toujours avec un épithélium cilié mais la musculature est plus développée que précédemment (pour la mastication).
- Le pharynx plissé : il est en forme de trompe dévaginable et rétractable, ce qui permet la capture de proies ou la succion de grosses proies.
Le rejet des excréments est précédé d’une entrée d’eau qui va être fortement expulsée alors qu’elle est chargée en excréments.
L’appareil excréteur
Les turbellariés sont les premiers animaux à posséder un appareil excréteur quoi que rudimentaire. Il est composé de deux canaux collecteurs latéraux longitudinaux. Ils collectent un réseau canaliculaire dense dans tout l’animal.
A l’extrémité de toutes les canalicules, on trouve une cellule excrétrice rudimentaire : protonéphridie, d’origine ectodermique. Ce type de cellule est aussi appelé « cellules à flamme ». Elles sont ciliées et c’est le battement de leurs cils qui pousse les déchets venant de l’hémolymphe dans des canaux collecteurs en direction des néphridiopores.
Cet appareil excréteur sert aussi à sortir l’eau du corps pour maintenir l’équilibre osmotique. Tout cela est vrai pour les formes d’eau douce qui ont un grand besoin de régulation osmotique. Ce n’est pas vrai pour les formes marines.
Le nombre de néphridiopores est différent selon le groupe de turbellariés (1, 2 ou plus).
L’appareil reproducteur et le développement.
Les turbellariés sont hermaphrodites.
L’appareil sexuel mâle est composé de testicules fragmentés en follicules testiculairesdeux spermiductes (ou canaux déférents) qui s’élargissent pour former les vésicules séminales à fonction de stockage des produits génitaux. drainés par
Ces deux vésicules confluent pour former un pénis tubulaire et musculeux. Ce dernier débouche dans l’atrium génital, commun aux deux sexes.
L’appareil sexuel femelle est constitué par deux ovaires en position antérieure, drainés par des oviductes (ou vitelloductes) où l’on trouve de nombreuses glandes vitellogènes qui produisent les cellules vitellines, cellules nourricières pour l’œuf.
Lors de la reproduction, les gamètes mâles et femelles confluent vers l’atrium génital où il existe un réceptacle séminal (ou « bourse copulatrice ») qui favorise le mélange des gamètes. Toutefois, il n’y a pas d’autofécondation car ce sont des hermaphrodites protandres (la maturité sexuelle mâle arrive avant la maturité sexuelle femelle).
Les œufs se retrouvent dans des cocons, par groupe de 4 à 6 où l’on trouve de nombreuses cellules vitellines.
La segmentation de l’embryon est de type spirale. Le développement est direct, sans stade intermédiaire larvaire.
 La multiplication asexuée.
La multiplication asexuée.
La multiplication asexuée peut se faire selon deux modalités :
- Par architomie : il y a division d’un individu en deux puis régénération des parties manquantes.
- Par paratomie : il y a d’abord régénération puis division.
C’est l’architomie qui est la plus souvent rencontrée. Les capacités à régénérer les parties manquantes s’observent chez certains groupes comme dans le cas d’un découpage transversal. Un découpage longitudinal donnera un individu avec deux régions antérieures.
 Systématique.
Systématique.
La systématique est basée sur la forme et sur la structure de l’appareil digestif. On distingue :
- L’ordre des Acèles (ou Acœles) : l’appareil digestif est réduit. On y trouve la bouche ainsi qu’une petite cavité buccale (il n’y a ni pharynx, ni intestin). Les formes de cet ordre sont essentiellement marines.
- L’ordre des Rhabdocèles : ils ont une bouche, un pharynx et un intestin rectiligne simple.
- L’ordre des Polyclades : L’intestin est très développé et ramifié. Les ramifications sont rayonnantes par rapport au centre de l’animal.
- L’ordre des Triclades : L’intestin est formé de trois branches (une antérieure et deux postérieures). De plus, chacune des branches est diverticulée.
Chez les triclades, on trouve les planaires au sens strict. On pourra distinguer trois sous-ordres en fonction de l’écologie des espèces :
- Les Terricoles sont terrestres.
- Les Maricoles sont marins.
- Les Paludicoles sont d’eau douce.
2 Les Trématodes.
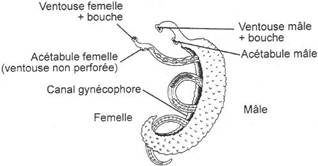
Les trématodes sont des parasites internes de vertébrés. Leur cycle fait intervenir deux ou plusieurs hôtes : ce sont des hétéroxènes. Ils ressemblent aux turbellariés. Les grandes différences sont dues au parasitisme :
- Il y a perte de la ciliature de l’épithélium (pas de déplacement).
- Il y a régression des organes des sens (en relation avec la perte de la locomotion) è On va parler d’une « évolution régressive » observée chez les trématodes.
- Il y a développement d’organes de fixation qui sont souvent une ventouse buccale antérieure et une ventouse ventrale ou « acétabulum ».
Les animaux à deux ventouses sont appelés « distomiens ».
Le développement est indirect avec de nombreuses formes larvaires asexuées à il y a un développement emboîté chez les trématodes.
Le tégument.
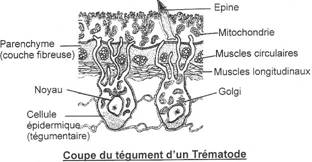
Le tégument des trématodes ressemble à celui des turbellariés. Ce tégument est recouvert par une cuticule avec des écailles ou des épines.
La présence d’une cuticule a deux principales conséquences : le développement ciliaire est impossible sur l’épithélium ; la croissance est alors discontinue et est réalisée par mues successives.
La cuticule a une lame basale. En dessous, on trouve un parenchyme et différents faisceaux musculaires (circulaires et longitudinaux). Sous cet ensemble, on trouve des cellules épidermiques qui peuvent émettre des diverticules jusqu’à la cuticule. Les cellules épidermiques n’ont pas de membrane cellulaire : c’est un tégument syncitial.
Système nerveux.
Le système nerveux est concentré en arrière de la ventouse buccale. Sa structure est proche de celle des turbellariés, avec des ganglions cérébroïdes reliés par une commissure. Des terminaisons nerveuses partent vers l’avant et des cordons nerveux partent vers l’arrière.
Différence : il y a absence d’organe des sens car les trématodes sont des parasites. Il leur reste quelques cellules sensorielles spécialisées.
L’appareil digestif.
La bouche est antérieure, terminale ou sub-terminal et s’ouvre au centre de la ventouse buccale.
L’intestin est formé par deux branches (cæcum digestif), très ramifiées où vient s’accumuler le sang ingéré.
Il n’y a pas d’anus.
L’appareil excréteur.
La structure de l’appareil excréteur est identique à celle des turbellariés (àprotonéphridien) avec un réseau caniculaire dense. Les canicules se réunissent en deux troncs principaux qui débouchent dans un réservoir terminal (« vessie ») qui s’ouvre à l’extérieur, par l’orifice excréteur (en position caudale).
L’appareil reproducteur.
Les trématodes sont hermaphrodites.
- L’appareil mâle :
Cet appareil est formé de deux testicules (ou follicules testiculaires) drainés par deux spermiductes qui convergent en une vésicule séminale de stockage. De cette vésicule se différencie un pénis copulateur qui fait saillie au niveau du gonophore (orifice mâle et femelle).
- L’appareil femelle :
L’appareil femelle est relativement complexe. Les fonctions vitellogènes et gamétogènes sont dissociées dans l’espace. Il gravite autour d’une chambre (l’ootype) où arrive l’oviducte, venant d’un ovaire ramifié. L’ootype reçoit un canal issu de la fusion des deux vitelloductes qui amènent les cellules vitellines venant des glandes vitellogènes (très ramifiées).
L’ootype reçoit aussi un utérus large et sinueux où les œufs commencent leur développement. Il y arrive en dernier un canal vertical (canal de Laurer) à fonction non définie.
La fécondation se fait par accolement réciproque.
L’œuf fécondé est entouré d’une coque et accompagné de nombreuses cellules vitellines. La reproduction s’effectue au niveau de l’hôte intermédiaire (pendant le cycle). Un œuf donnera alors de nombreux individus.
Le cycle parasitaire.
Ce cycle comporte de nombreux stades : on parle alors de « développement emboîté ». Les diverses formes larvaires viennent du cycle théorique des trématodes (plusieurs stades larvaires avant le stade adulte).
L’œuf a une coque qui possède un opercule apical (et des cellules vitellines). L’éclosion donne une petite larve ciliée (larve miracidium) avec un cerveau rudimentaire (glandes céphaliques), deux protonéphridies et un grand nombre d’amas de cellules germinales. C’est un stade libre qui peut infecter un animal (mollusque). Il y a alors enkystement dans la glande digestive du mollusque : on passe au stade sporocyste. Cette dernière forme a la forme d’un sac à paroi souple avec des amas de cellules germinales qui vont se multiplier et donner naissance à un troisième stade larvaire (la rédie). Un sporocyste contient de nombreuses rédies. Ces rédies ont deux devenirs possibles :
- donner des cercaires, ce qui est le 4ème stade larvaire,
- ou donner des rédies filles, ce qui permet une multiplication du nombre de rédies.
Le stade cercaire possède, comme l’adulte, deux ventouses (dont une antérieure buccale), un cerveau rudimentaire et un appareil digestif à deux branches.
A ce stade, il y a sortie de l’hôte intermédiaire puis enkystement qui donne la forme métacercaire : c’est le stade infectant de l’hôte définitif, qui donnera l’adulte.
Un œuf donnera de nombreux individus grâce aux diverses multiplications larvaires asexuées du cycle.
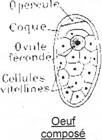
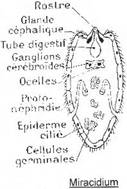
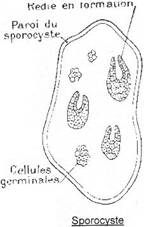
Exemple : la petite douve du foie, Dicrocœlium dendriticum.
Cette douve vit dans les canaux biliaires du mouton. L’adulte mesure environ 10 millimètres de long et 2 à 3 millimètres de large. Il existe deux hôtes intermédiaires :
- un gastéropode terrestre (Hélix)
- et une fourmi (Formica).
Les œufs sont rejetés avec les excréments du mouton, mangés par un gastéropode (avec une coquille blanche à bandes brunes). A l’automne, il y a éclosion des œufs (à larve miracidium mobile). Ces larves se dirigent vers la glande digestive où elles s’enkystent en sporocystes. Ces derniers donnent des cercaires (il n’y a pas de stade rédie). Les cercaires sortent du gastéropode pour être avalés par une fourmi. Il va y avoir enkystement dans la cavité générale des fourmis : c’est le stade métacercaire. Les fourmis vont alors avoir un comportement parasité et grimper sur des brins d’herbe pour être avalées par un mouton.
La petite douve peut infecter l’Homme par ingestion de foie de mouton mal cuit, mais c’est une infestation accidentelle.
Exemple : La grande Douve du foie, Fasciola hepatica.
L’adulte se trouve dans le foie de mouton. L’hôte intermédiaire est un gastéropodeinfesté par le stade miracidium (qui est libre et nageur dans le milieu aquatique). aquatique (Limnée)
La larve miracidium, par enkystement en sporocyste, donne des rédies. Selon les conditions de température, les rédies ont deux devenirs possibles : si l’eau a une température supérieure à 25°C, elles redonnent des rédies (rédies filles) ; si la température est inférieure à 25°C, on passe au stade cercaire.
Le stade cercaire se fixe et s’enkyste en métacercaire sur les plantes aquatiques comme le cresson. Si l’Homme consomme des végétaux souillés, il peut être contaminé et être atteint de distomatose. Toutefois, l’Homme n’est pas indispensable au cycle de Fasciola.
Exemple : la douve pulmonaire.
Cette douve provoque des bronchites sanglantes ressemblant à la tuberculose ou à la pneumonie. Son cycle fait intervenir : l’Homme, un mollusque (Melania) et un crabePotamon). (
Le mollusque est le lieu de développement de stades larvaires, jusqu’au stade cercaire. Le crabe permet le passage du stade cercaire au stade métacercaire.
Exemple : Schisostome (bilharziose).
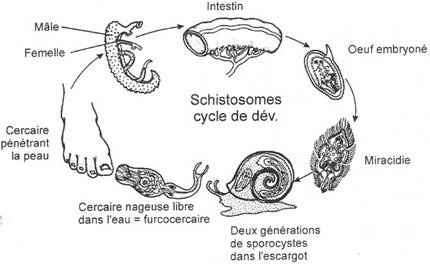
Ici, les sexes sont séparés. Les adultes vont dans le système veineux des mammifères. Il y a dimorphisme sexuel important : le mâle possède un canal gynécophore, une gouttière dans laquelle la femelle s’installe pendant l’accouplement.
L’hôte intermédiaire est un mollusque où la larve miracidium donne un sporocyste puis le stade cercaire (nageur) qui sera libéré. Les furocercaires vont contaminer l’Homme en pénétrant par la peau lors d’une baignade (15 minutes sont nécessaires pour qu’un furocercaire rentre complètement).
Dans ce cycle, il y a absence des stades rédies et métacercaires : c’est une diminution des formes larvaires. Le cycle devient plus rapide avec moins d’évènements hasardeux.
3 Les Monogènes (= Polystomiens).
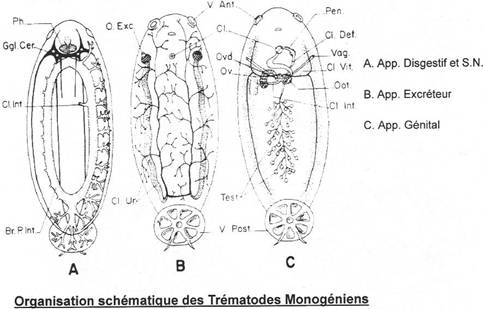
Les monogènes sont des parasites monoxènes et monogéniens (une seule forme larvaire). Selon les espèces, on peut avoir des parasites externes de poissons, de crustacés, de céphalopodes ou de batraciens. Généralement, ce sont des parasites provoquant peu de dommages. Ils sont surtout dangereux pour les élevages intensifs.
Morphologie.
La bouche est antérieure ou ventrale. Il y a plusieurs ventouses : la ventouse antérieure est appelée « cotylophore » ; les autres ventouses sont postérieures, accompagnées de crochets.
On note la présence d’un canal génito-intestinal servant à évacuer le trop plein de cellules vitellines.
Exemple : Polystoma integerrinum.
C’est un parasite de la grenouille rousse (Rana temporaria). Il y a un synchronisme entre les cycles biologiques de ces deux espèces (certaines phases du cycle sont conjointes). Au moment de la reproduction (ponte) de la grenouille, les Polystoma adultes pondent des œufs dans les urines (et sont évacués en milieu aquatique).
Ces œufs donnent une larve ciliée nageuse ; un oncomiracidium qui peut se fixer sur les têtards de grenouille rousse.
Si le têtard a plus de 10 jours, la larve gagne l’appareil digestif du têtard et va atteindre la vessie pour donner un adulte. L’adulte est apte à se reproduire au bout de trois ans, comme les batraciens.
Si la larve rencontre un têtard de moins de 7 à 10 jours, la suite du cycle parasitaire ne peut avoir lieu : pas d’infestation.
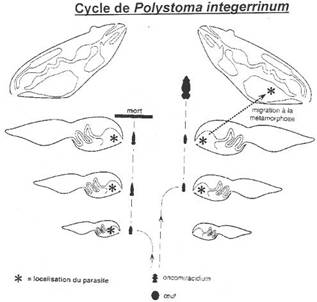
Exemple : Diplozoon paradoxum.
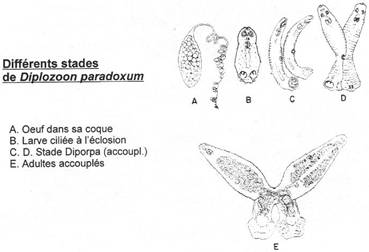
L’adulte se fixe sur les branchies d’un poisson d’eau douce (Cyprinidés). Le parasite présente une espèce constituée de la fusion de deux individus où chacun est incapable de survivre isolément.
Les larves sont aspirées par les poissons et se fixent sur les branchies. Elles y subissent une métamorphose qui différencie une ventouse circulaire dans la partie médiane ventraleet un petit bouton sur la face dorsale, formé par un soulèvement de la cuticule. Ce sont des larves diporpas qui se nourrissent du sang du poisson pendant plusieurs semaines. La suite du développement se fait si deux larves entrent en contact l’une avec l’autre. L’accolement se fait par la ventouse qui attrape le bouton, chez l’un comme chez l’autre. (de la larve)
Si l’accolement se fait, il y a fusion de ces larves au niveau de leurs points d’attache è c’est une anastomose entre les deux intestins : la nourriture ingérée par l’une est alors utilisable par l’autre. Il y a aussi fusion des conduits sexuels mâles et femelles.
è Les deux individus sont définitivement accouplés. La fusion met en connexion les canaux déférents de l’un avec le vagin de l’autre : c’est une fécondation réciproque croisée.
4 Les Cestodes (classe).
Les cestodes sont des parasites du tube digestif de vertébrés supérieurs. Leur corps est aplati, segmenté, plus ou moins rubané. Ce sont des animaux généralement longs : 12 à 20 centimètres pour les plus grands. Ils n’ont ni tube digestif ni appareil respiratoire.
Morphologie.
Le corps est divisé en trois parties :
- Région antérieure. C’est le scolex ; il porte le dispositif de fixation : ventouse et/ou crochets en couronne.
- Zone de prolifération ou cou. C’est à ce niveau que se forment les segments composant le corps de l’animal.
- Le strobile. Il occupe 9/10ème du corps. Il est formé d’une succession de segments (les proglottis).
Le tégument.
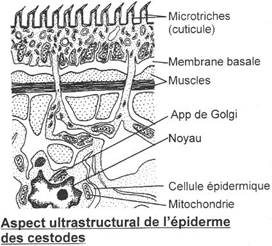
Le tégument est composé d’une cuticule épaisse permettant une protection face aux enzymes digestives de l’hôte. Cette cuticule est recouverte par des microtriches (épines). Elle repose sur la membrane basale. En dessous, on observe deux couches musculaires : une couche musculaire circulaire externe et une couche musculaire longitudinale interne. Sous les muscles, on trouve des cellules épidermiques profondes gardant des relations avec les couches superficielles du tégument.
Le système nerveux.
Le système nerveux est situé dans le scolex et est constitué de deux ganglions cérébroïdes présentant des prolongements antérieurs et postérieurs.
Les prolongements antérieurs s’unissent par une commissure circulaire où se situent deux ganglions dorsaux et deux ventraux.
De chaque ganglion part un cordon nerveux, à l’intérieur du strobile.
Vers l’arrière, on trouve une commissure avec deux ganglions latéraux d’où partent deux cordons latéraux dans le strobile.
A l’extrémité postérieure des proglottis, on a une liaison (commissure) qui relie les six cordons nerveux avec les deux ganglions.
Il n’y a pas d’organe sensoriel différencié mais de nombreuses terminaisons nerveuses aboutissent à la cuticule.
L’appareil excréteur.
Cet appareil excréteur est composé de 4 canaux excréteurs tout le long du strobile : deux ventraux et deux latéraux.
Dans la région postérieure de chaque proglottis, on a une anastomose qui réunie les canaux ventraux. Ils débouchent à l’extérieur, au niveau du dernier proglottis du strobile alors que les canaux dorsaux s’atrophient avant d’atteindre l’extrémité du strobile.
L’appareil reproducteur.
L’appareil reproducteur rappelle celui des trématodes ; il y a hermaphrodisme. Un pore génital est commun aux deux sexes : c’est l’atrium génital. L’hermaphrodisme est protandre. On trouve les appareils mâle et femelle dans le même proglottis.
Les proglottis antérieurs porteront l’appareil mâle.
Les proglottis moyens auront un appareil mâle et un appareil femelle bien développé.
Les proglottis âgés auront un appareil mâle dégénéré et un développement important de l’utérus qui se ramifie et va contenir de nombreux œufs en développement (»5000 œufs par utérus).
Classification.
La classification des cestodes est réalisée à partir de la structure du scolex, de l’appareil génital et sur les particularités du cycle parasitaire. On distingue deux sous-classes :
- Les Cestodaires : ils n’ont pas de scolex.
- Les Eucestodes : ils possèdent un scolex. Cette sous-classe regroupe deux ordres :
· L’ordre des Cyclophyllidiens. Le scolex possède quatre ventouses avec des fibres musculaires radiaires.
· L’ordre des Pseudophyllidiens. Le scolex présente deux dépressions (pseudobothridies), une dorsale et une ventrale. La musculature est peu différenciée.
Développement.
Les cestodes sont dans le tube digestif de leur hôte définitif et s’y nourrissent par osmoseosmotrophes ». Ils n’ont pas d’appareil respiratoire car ils ont une vie en anaérobiose. et n’ont donc pas de tube digestif : ce sont des «
Pour pouvoir réaliser tout leur cycle de développement, ils se sont glissés dans la chaîne alimentaire. Ils ont un grand pouvoir reproducteur ; ils sont composés d’un grand nombre de segments (proglottis) formés par strobilation, ce qui est assimilable à une reproduction asexuée (fournie par des entités reproductrices complètes).

Les proglottis présentent un hermaphrodisme protandre
La fécondation est réalisée quand un repli de l’animal met en relation des proglottis de sexe différent. A la suite de la fécondation, les cucurbitains donnent des millions d’œufs. Toutefois, tous les cestodes ne sont pas aussi féconds : de petites espèces compensent leur faible nombre d’œufs par une multiplication asexuée à l’état larvaire.
Un œuf fécondé donnera par divisions, un embryon entouré de 3 ou 4 blastomères. Le tout est entouré d’une coque. L’embryon développe trois paires de crochets è c’est la larve hexacanthe enveloppée par une coque (à embryonophore). Embryonophore + embryon hexacanthe = oncosphère. Cette dernière peut être aquatique et libre et elle sera appelée larve coracidium.
Le cycle de développement ne se poursuit que si le stade oncosphère est ingéré. Les sucs digestifs détruisent la coque, cassent l’embryonophore et libèrent l’embryon. Ce dernier se fixe par les crochets, perfore l’épithélium et se dirige vers la musculature par la voie sanguine. Une fois dans les muscles, il y a perte des crochets. Il se forme alors une invagination qui donnera le futur scolex è c’est un cysticerque.
Le stade cysticerque est infestant. S’il rencontre un deuxième hôte, il se transforme en procercoïde puis en plérocercoïde.
Le cysticerque est une vésicule sphérique, creuse, avec un scolex invaginé. Pendant sa phase d’attente, le cysticerque s’enkyste dans les muscles.
La larve procercoïde est une larve allongée, dépourvue de cavité interne. Son corps présente une constriction délimitant deux parties : à l’avant, on a la future larve et la partie postérieure dégénère.
La larve plérocercoïde possède le scolex du vers adulte et un début de segmentation. Le corps est allongé (plusieurs millimètres de long), d’une couleur blanchâtre qui est due à la présence de particules calcaires dans le parenchyme.
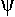 Cycles.
Cycles.
· Tænia saginata (Cyclophyllidiens).
Ce vers mesure de 4 à 12 mètres de long et sa durée de vie atteint 30 ans et même plus. Son scolex est dépourvu de crochets mais il possède quatre ventouses.
Les pores génitaux sont disposés irrégulièrement à droite ou à gauche le long du strobile. Les cucurbitains sont libérés au rythme d’une quinzaine par jour, de manière passive ou active (ils peuvent forcer le sphincter anal du sujet parasité). Les cucurbitains ont un utérus très ramifié (20 à 35 digitations chez le saginata). Dans chaque cucurbitain, on peut trouver jusqu’à 80 000 œufs.
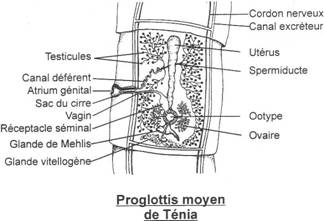
Les stades cysticerques se rencontrent dans la musculature des bovidés (c’est alors le Cysticercus bovis). L’infestation est possible par ingestion de viande de bœuf crue ou pas assez cuite.
· Tænia solium.
Ce vers peut mesurer jusqu’à trois mètres de long (soit » 1000 proglottis). Le scolex possède une double couronne de crochets. Les pores génitaux alternent régulièrement le long du strobile. La fécondation se fait par un repli du strobile sur lui-même.
Les cucurbitains se détachent par groupe de 5 ou 10 de façon passive. L’utérus comprend entre 10 et 12 digitations.
Les formes larvaires sont dans la musculature du porc (cœur et langue). Les cysticerques sont appelés « Cysticercus cellulosae ».
L’infestation est réalisée en mangeant de la viande de porc crue ou pas assez cuite. Le vers devient adulte en trois mois. L’Homme peut s’infester lui-même en ingérant des oncosphères par l’intermédiaire de mains souillées ou de remontée de cucurbitains dans l’estomac. è C’est la cysticercose : cette maladie peut atteindre le cerveau ainsi que les organes sensoriels tels que les yeux.
· Bothriocéphale : Diphyllobothrium latum.
Ce vers peut atteindre 2 à 8 mètres et même parfois, 20 mètres. Le scolex est ovoïde et porte deux pseudobothridies (c’est un pseudophyllidé). On y trouve entre 3000 et 4000 anneaux avec un pore génital en position médiane. Les adultes se trouvent dans l’intestin de l’Homme, du chat, du chien, du renard. Les oncosphères sont éliminées avec les selles.
La larve est ciliée, nageuse à larve coracidium. Ce stade larvaire va être avalé par un crustacé du genre copépode dans l’intestin duquel il perd sa ciliature et puis, pénètre dans la cavité générale. C’est alors une larve procercoïde.
Le développement s’arrête et la larve vit aussi longtemps que le crustacé. Si ce dernier est capturé par des poissons carnassiers, le cycle continue. Si le copépode est ingéré par un poisson non carnassier, le cycle est bloqué et ne pourra reprendre que si la larve se retrouve finalement dans un poisson carnassier.
Dans l’estomac du poisson carnassier, les larves entrent dans la paroi et vont se loger dans la musculature (surtout dans les flancs). Du stade procercoïde, on passe ou stade plérocercoïde qui est infestant pour l’Homme. Si cette dernière larve est mangée par un hôte définitif, elle donnera le stade adulte assez rapidement.
Dans certains cas, le poisson carnassier peut être mangé par un autre poisson : les larves pleurocercoïdes ne restent alors pas dans l’estomac mais partent dans les viscères du dernier poisson. Là, elles pourront survivre pendant plusieurs années, sans grandir ni se différencier. Le cycle continue si ce poisson est mangé par un poisson carnassier…
· La Ligule intestinale : Ligula intestinalis.
La ligule adulte se trouve dans l’intestin d’oiseaux aquatiques piscivores (comme les cormorans ou les canards). Le premier hôte intermédiaire est un crustacé copépode. Le deuxième hôte intermédiaire est un poisson cyprinidé. C’est dans la cavité abdominale de ces poissons que les larves se rencontrent. Elles peuvent atteindre plus de 25 centimètres de long et il peut y avoir plusieurs larves dans un même poisson. Il va se produire une déformation importante de la partie ventrale : le corps s’alourdie et la capacité de nage s’en trouve diminuée. Ces poissons deviennent plus faciles à attraper par les oiseaux.
Ce parasitisme entraîne une stérilisation des poissons car il n’y a plus d’espace libre pour le développement des gonades.
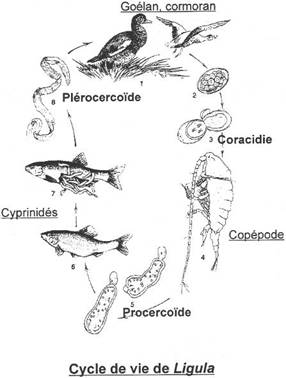
· Tænia multiceps.
Ce tænia vit dans l’intestin grêle. Sa forme larvaire est localisée dans le cerveau de mouton (c’est la forme cénure) dont la taille est celle d’un œuf de poule (6 à 8 centimètres). La présence de cette larve provoque des troubles de la mobilité : c’est la maladie du tournis.
C’est sur la forme larvaire que vont bourgeonner de nombreux scolex, au niveau de la membrane proligère. La formation de nombreux scolex s’apparente à une multiplication asexuée.
· Tænia échinocoque : Echinococcus granulus.
On trouve ce vers chez les canidés. Il peut mesurer 3 à 4 centimètres de long ; il possède une double couronne de crochets et 3 à 5 proglottis pour le strobile. Les deux premiers proglottis sont en cours de maturation ; le troisième possède des organes génitaux formés ; les deux derniers contiennent des œufs fécondés.
Les cucurbitains renferment chacun 300 à 800 œufs. Ces derniers sont libérés avec les excréments, puis ingérés par des herbivores. L’Homme peut être contaminé accidentellement par l’intermédiaire de crudités souillées ou bien, s’il touche un chien ayant portant le parasite (prurit anal).
Le vers traverse la paroi intestinale et rejoint le foie de l’herbivore et là, se différencie en une boule de 20 centimètres de diamètre : le stade hydatide. Ce stade se développe lentement : 16 mois chez les ovins et les bovins ; 18 mois à 30 ans chez l’Homme. Dans l’hydatide, il y a un bourgeonnement de scolex à partir de la membrane interne (proligère) : on peut arriver à 400 000 scolex par centimètre cube. Chez l’Homme, il y a besoin d’une intervention chirurgicale. Les plus grands foyers de développement sont les lieux d’élevage intensif de mouton.
· Echinococcus multilocularis (Echinococcus alveolaris).
Ce vers est un parasite du renard (hôte définitif). L’hôte intermédiaire est le campagnol. L’Homme peut être hôte intermédiaire s’il consomme des baies sauvages souillées. Il existe une forme hydatide (dans le foie) qui provoque des nécroses hépatiques très graves.
Chez l’Homme, cette maladie est l’échinococcose alvéolaire ; 200 cas étaient recensés en France, vers 1980.
· Diphylidium cannium (cyclophyllidés).
C’est un vers dont la taille varie entre 50 et 80 centimètres. L’adulte est dans l’intestin de chien. L’œuf se développe quand il est avalé par des larves de puces.
La puce subit des mues ou des métamorphoses et le chien se contamine en ingérant des puces adultes.
II Les Pseudocœlomates.
Les pseudocœlomates ont leurs organes situés dans une cavité corporelle qui a des avantages réels. Celle-ci est appelée cavité viscérale ou primaire. Toutefois, le mésoderme ne participe que partiellement à la délimitation de cette cavité (du coté externe uniquement).
La cavité renferme quelques cellules amiboïdes. Le tube digestif et les gonades sont suspendus dans cette cavité.
On définit sept embranchements dont trois qui seront étudiés en cours : les Nématodes, les Nématomorphes et les Rotifères. Anciennement, les nématodes et les nématomorphes composaient le groupe unique des némathelminthes.
Les pseudocœlomates ont tous un plan d’organisation et des caractères semblables :
- Ils sont tous plus ou moins vermiformes.
- Leur corps est recouvert d’une cuticule.
- La paroi du corps ne possède pas de couches musculaires circulaires et transversales.
- La paroi du tube digestif est généralement simple (cellules épithéliales sur une couche) avec parfois une couche musculaire autour de la paroi.
- Il n’y a ni système respiratoire ni système circulatoire.
- Le système excréteur est le plus souvent protonéphridien.
- Les pseudocœlomates présentent un cloaque où débouchent les systèmes digestif, urinaire et génital.
Toutefois, les nématodes femelles ont un gonopore séparé.
La plupart des pseudocœlomates présente un nombre défini de cellules dans chaque organe. à C’est l’eutélie. La croissance se fait grâce à l’augmentation du volume cellulaire et non du nombre de cellules. On pourra fréquemment observer des cellules géantes.
Toutefois, cette tendance limite les possibilités de régénération. En fait, cette tendance a été étudiée chez Cœnorhobditis elegans (génétiquement très connu, comme la drosophile) et il a été montré un même complexe de gènes homéotiques que chez la drosophile, la souris et l’Homme (HOM/HOX).
A Les Nématodes (semblables à un fil).
Chez les nématodes, on observe l’absence totale de cellule flagellée ou ciliée (même chez les spermatozoïdes). Les spermatozoïdes sont ronds ou coniques et se déplacent grâce à des mouvements amiboïdes.
Il existe plusieurs dizaines de milliers d’espèces (80 000) mais elles sont difficiles à identifier. Les nématodes sont souvent parasites de végétaux ou d’animaux (dont l’Homme) et provoquent des nématodoses (dangereuses).
D’un point de vue écologique, on les trouve sous toutes les latitudes (des glaciers jusqu’aux sources thermales) et ils présentent une grande uniformité structurale.
1 Morphologie
Les nématodes sont des vers blancs à section circulaire, effilés aux deux extrémités Les mâles ont une taille comprise entre 12 et 15 centimètres alors que les femelles sont bien plus grandes. La bouche s’ouvre à l’extrémité antérieure et est entourée de trois lèvresune dorsale et deux ventrales) portant des soies céphaliques à rôle sensoriel. (
La femelle a une extrémité postérieure qui porte l’anus. Le mâle a une extrémité postérieure enroulée ventralement, en crosse, et qui porte, en position sub-terminale, un orifice sub-cloacal où l’on trouve des spicules sexuels copulateurs.
La femelle présente une constriction annulaire dans le tiers antérieur du corps et c’est à ce niveau que s’ouvre l’orifice génital ventral.
Lors de l’accouplement, le mâle enroule sa crosse autour de la femelle, dans le tiers antérieur.
2 Tégument.
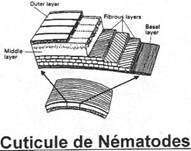
L’épiderme est recouvert par une cuticule épaisse, imperméable, pouvant avoir jusqu’à neuf couches cellulaires (dont trois couches de fibres obliques croisées). Ces fibres permettent de résister à la très forte pression interne car ils n’ont pas de musculature interne. Cette cuticule limite aussi les pertes par évaporation.
Sous la cuticule se trouve l’épiderme avec deux épaississements longitudinaux (un dorsal et un ventral) où sont localisés les cordons nerveux (un dorsal et un ventral). Latéralementdeux gouttières au niveau desquelles on a les canaux excréteurs. (à gauche et à droite) on trouve
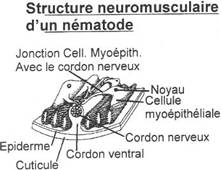
Le mésoderme est représenté par des cellules myo-épithéliales. Elles sont implantées dans le feuillet ectodermique externe ; elles vont définir la paroi externe de la cavité primaire (ou cavité viscérale) è la somatopleure.
Ces cellules sont sur une assise et forment quatre champs musculaires (deux dorsaux et deux ventraux). Ces champs sont délimités par quatre cordons (un dorsal, un ventral, un à droite et un à gauche). Ces cellules différencient des myofibrilles du côté de l’épiderme : c’est un dispositif original car d’habitude, ce sont les nerfs qui envoient des fibrilles vers les organes. La contraction de ces cellules donnera uniquement des enroulements.
3 Les organes des sens.
Il existe des organes classiques mais aussi :
- Des Amphides : invaginations cuticulaires dans la partie antérieure à rôle chimiorécepteur.
- Les Phasmides : ce sont des organes sensoriels paires, situés latéralement en position post-annulaire.
è Les organes sensoriels permettent de distinguer deux sous-groupes.
4 Le système nerveux.
Le système nerveux des nématodes est rudimentaire, acculé ou inclus dans l’épiderme. Il est composé de centres nerveux localisés au niveau du collier péri-œsophagien. Ce collier émet des nerfs en direction des lèvres buccales et va émettre des troncs nerveux moteurs ou sensitifs. On distingue six nerfs dans la région antérieure et huit troncs nerveux (6 moteurs et 2 sensitifs) qui partent vers la partie postérieure.
5 L’appareil excréteur.
L’appareil excréteur ne porte pas de protonéphridie. On a deux types distincts :
- Chez les nématodes primitifs, on a une ou deux cellules géantes (les cellules Renette) situées ventralement à la jonction œsophage/intestin. Chaque cellule possède un pore excréteur en position antérieure.
- Chez les formes plus évoluées, les cellules géantes s’atrophient et il y a formation de deux canaux excréteurs qui courent le long du corps ; un seul pore excréteur est présent en position antérieure.
6 L’appareil digestif.
L’appareil digestif est pratiquement rectiligne et comporte cinq éléments principaux :
- La bouche (en position terminale avant) est entourée par trois lèvres hémisphériques (une dorsale et deux ventrales).
- La cavité (ou capsule) buccale est tapissée par une mince couche cuticulaire ; l’organisation est fonction du comportement alimentaire des différentes formes. Cette capsule buccale peut être armée de dents, denticules ou râpes. Chez les nématodes marins les plus primitifs, quatre formes existent :
· Forme simple sans armature : elle permet l’absorption de nourriture liquide.
· Forme conique sans armature : on la trouve chez les nématodes microphages.
· Forme à armature réduite (denticules ou râpes) : elle est présente chez les mangeurs d’algues ou chez certaines formes mégalophages (qui mangent de tout).
· Forme à armature développée : existe chez les nématodes prédateurs.
- Le pharynx est musculeux à parois épaisses, tapissé d’une couche cuticulaire. C’est à ce niveau que l’on trouve des glandes à fonction anti-coagulantes (pour les formes parasites de vertébrés).
- L’intestin est droit et mince (une seule couche cellulaire endodermique).
- Le rectum est d’origine ectodermique car il est tapissé d’une couche cuticulaire. Les femelles ont un rectum terminé par un anus qui s’ouvre en position sub-terminale. Chez le mâle, le rectum débouche dans un cloaque où l’on trouve les spicules copulateurs. C’est ici que s’ouvrent les canaux déférents.
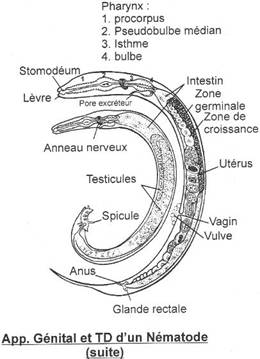
Les nématodes ingèrent souvent de la nourriture liquide, et, pour se faire, il existe chez eux, un dispositif pouvant contrebalancer leur pression interne importante. Ce dispositif est présent au niveau du pharynx et terminé par une valve. Au milieu du pharynx, on a une sorte de pompe pharyngienne :
- D’abord, la partie antérieure se dilate : il y a aspiration de liquide ; la valve terminale est fermée.
- Ensuite, le pharynx se contracte : la partie antérieure va avoir un diamètre plus petit ; la valve postérieure s’ouvre et le liquide passe par cette valve.
7 L’appareil génital.
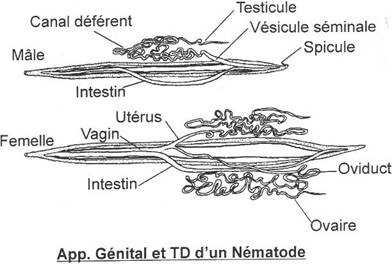
Les nématodes sont des animaux gonochoriques.
L’appareil mâle :
Cet appareil est constitué d’un testicule tubulaire, filiforme, très long (1 mètre ou plus). Il est entouré sur lui-même et autour de l’intestin. Ce testicule se renfle dans la portion terminale et forme la vésicule séminale qui stocke les spermatozoïdes. Cette vésicule débouche dans le cloaque.
L’appareil femelle :
L’appareil femelle est composé de deux ovaires tubulaires filiformes (un à deux mètres). Ces ovaires se transforment ensuite en oviductes puis en utérus bien développés. Ils vont se rejoindre pour donner le vagin qui est beaucoup plus grêle. Il aboutit à l’orifice génital femelle (dans le tiers de la partie antérieure, ventral).
Les femelles fécondées pondent de 70 000 à 240 000 œufs par jour pendant 12 à 18 mois.
La fécondation est toujours croisée et interne grâce aux stylets copulateurs du mâle.
Les œufs sont pondus à différents stades selon l’espèce. La ponte peut donner :
- Des œufs non segmentés.
- Des œufs à segmentation débutée.
- Des œufs à embryon complètement formé.
Il existe une exception au gonochorisme des nématodes chez le genre Rhabdite : l’hermaphrodisme est protandre, avec autofécondation.
La cuticule externe oblige l’accroissement à se faire par mues successives (en général, il y a quatre mues dans la vie d’un animal).
8 Les grands groupes de nématodes.
On distingue deux principaux groupes :
- Les Aphasmidiens : ils n’ont pas de phasmide et sont parasites.
- Les Phasmidiens : ils possèdent deux phasmides et sont libres, vivant dans la terre ou dans la matière putréfiée ; sinon, ils peuvent être parasites de végétaux ou d’animaux.
9 Ecologie et éthologie des formes libres.
Les formes libres représentent la moitié des nématodes. On les rencontre dans la plupart des habitats (eau douce, eau de mer, sol humide, mousse, matière organique en décomposition).
Souvent, les nématodes pullulent :
- Dans les fruits pourris des vergers : on trouve plusieurs milliers d’individus d’espèces différentes.
- Dans les milieux hostiles au développement de la vie. Exemple : l’anguille du vinaigre (Anguillula) se développe dans le vinaigre, avec 6 à 7% d’acide acétique pur. Elle se nourrit du cadavre des bactéries acétiques, responsables de la fermentation. Elle supporte des pH allant de 2,5 à 11,5.
10 Ecologie et éthologie des formes parasites.
Généralement, en cas de parasitisme, il y a des modifications structurales importantes et présence d’une reproduction asexuée possible à l’état larvaire.
Ces modifications ne se voient pas chez tous les nématodes. Le développement des formes libres ou parasites est identique. Les parasites vont s’attaquer aux végétaux ou aux animaux.
a Les parasites de végétaux.
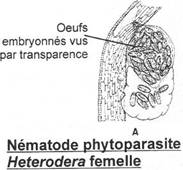
Ces nématodes sont des destructeurs redoutables des végétaux cultivés. Ils provoquent l’interruption de la croissance de la plante, une castration (par suppression des fruits et des graines), entraînent l’apparition de galles spécifiques. Les galles sont des kystes sur les tiges ou sur les racines où vivent les femelles qui se nourrissent de cellules géantes, produites par la plante parasitée.
b Les parasites d’animaux.
Exemple 1 : Enterobius vermicularis.
Ce parasite est responsable de l’oxyurose.
Les adultes vivent et s’accouplent dans le cæcum digestif de l’Homme, pouvant provoquer l’appendicite. Après accouplement, les femelles vont dans le rectum, s’y fixent et pondent. Il y a alors démangeaisons ou prurit anal. Les œufs (de 50 à 60µm) restent collés dans l’anus. Ils donnent les embryons qui sont directement infestant.
L’enfant se contamine en portant les mains (souillées) à la bouche (c’est une auto-infestation ou parasitose des mains sales), ou en inspirant des poussières contenant des œufs.
Les œufs, une fois dans le tube digestif vont subir l’action des sucs digestifs qui vont casser la coque et ainsi, libérer les larves qui passent au stade adulte en une trentaine de jours. Les adultes se nourrissent des bactéries du tube digestif et ne sont pathogènes que lorsque leur population augmente trop fortement et provoque des occlusions du tube digestif.
Exemple 2 : Trichurus trichuria (monoxène).
Ces vers sont des parasites des cæcums digestifs. Ils ont une partie antérieure effilée, implantée dans la muqueuse digestive des mammifères (la partie postérieure est libre). Ce sont des hématophages. Une centaine d’individus consomment 50 millilitres de sang par jour.
Les œufs pondus ne donnent pas directement des larves (en restant à l’intérieur). Pour la suite de leur développement, un séjour de trois semaines à l’extérieur est obligatoire.
La durée de vie de l’adulte est de cinq ans. Les femelles donnent 10 000 œufs par jour et l’infestation se fait par l’intermédiaire de boissons et de légumes mal lavés.
Les œufs donnent des adultes en un mois.
Exemple 3 : Ascaris.
Les larves d’ascaris font d’importants déplacements dans l’organisme parasité : migration dans l’hôte (à cycle avec migration).
Les lombricoïdes provoquent l’ascaridiose humaine.
Les adultes s’accouplent et pondent dès l’intestin grêle. La femelle donne 240 000 œufs par jour. Ces derniers seront évacués avec les excréments et donnent des larves infestantes au bout de 30 à 40 jours si la température externe est voisine de 30 degrés, avec des sols humides et ombragés.
L’infestation est réalisée par ingestion d’aliments souillés. Après la digestion de la coque, la larve va migrer dans l’organisme. Les larves traversent d’abord l’épithélium digestif avant d’atteindre le foie par voie sanguine. Elles y restent pendant trois à quatre jours puis, gagnent le cœur droit par la circulation veineuse puis rejoignent les poumons par les artères pulmonaires et y restent environ une semaine. Elles vont y subir deux mues. Elles remontent ensuite les voies respiratoires (bronchioles, bronches, trachée) et arrivent au niveau du carrefour bucco-pharyngé et prennent alors la voie digestive et rejoignent le grêle où elles deviendront adultes. La migration dure environ 15 jours. Après le grêle, les larves passent au gros intestin, puis au rectum où elles sont matures sexuellement.
L’homme est le seul réservoir de ce parasite et cette maladie touche environ ¼ de la population mondiale, surtout dans les pays pauvres.
Les symptômes observés sont : des occlusions intestinales, des invaginations intestinalesà hernies), des péritonites aiguës et des dégâts dans le foie (pancréatites). De plus, ces ascaris rejettent des substances toxiques pour le système nerveux de l’Homme. D’autres vertébrés or l’Homme peuvent être parasités par pratiquement la même espèce (cheval, porc, chien, chat). (et des étranglements de ce tube digestif
Exemple 4 : Trichinella spiralis (hétéroxène).
On trouve ce parasite dans l’intestin grêle de nombreux animaux (chien, chat, rat, porc, sanglier, phacochère, Homme). Les femelles pénètrent dans la muqueuse intestinale et y pondent deux larves par heure pendant les vingt-huit jours de leur vie.
Les femelles sont vivipares et donnent des larves qui gagnent le cœur droit par le liquide lymphatique puis, les poumons, passent dans le cœur gauche et seront distribuées dans tout l’organisme par la circulation aortique. Ces larves vont se fixer dans les muscles striés et s’enkyster pendant 12 à 18 mois. Les kystes se calcifient et sont infestant. Les Hommes se contaminent en consommant de la viande mal cuite.
Chez les rats, l’infestation est propagée par le cannibalisme caractérisant cette espèce : l’hôte intermédiaire et l’hôte définitif peuvent être de la même espèce. Dans ce cas, on parle alors de cycle auto-hétéroxène.
Exemple 5 : Wuchereria bankrofti (Filaire de Bancroft).
Les adultes de ce parasite vivent dans les vaisseaux lymphatiques. Le mâle fait environ 4 centimètres et la femelle mesure aux alentours de 8 centimètres. Les larves circulent dans le liquide lymphatique et migrent périodiquement dans les vaisseaux sanguins périphériques selon un rythme nycthéméral (jour/nuit) correspondant à la sortie de l’insecte vecteur (Anophèle : moustique). Grâce aux piqûres, les moustiques pompent le sang de vertébrés alors que les larves se concentrent sur les pièces buccales du moustique, Elles seront transmises à la victime suivante par une nouvelle piqûre. Les larves deviennent adultes en trois mois. Les parasites sont surtout dans les zones tropicales (Afrique, Amérique du sud, Asie).
Ce parasite provoque l’éléphantiasis : les membres inférieurs deviennent énormes à cause de la présence du parasite dans le système lymphatique de ces membres. Le scrotum peut aussi être atteint.
B Les nématomorphes.
Les nématomorphes sont des vers longs (environ 1,5 mètre) qui ressemblent aux nématodes. Leur intestin est réduit, les larves sont parasites alors que les adultes sont aquatiques libres.
La vie adulte est brève et ces adultes ne se nourrissent pas : l’anus et la bouche sont non-fonctionnels. La larve va donc se nourrir par osmose, au travers du tégument.
Le système nerveux est intra-épidermique ; canaux excréteurs et protonéphridies sont absents ; on note la présence de cellules myo-épithéliales sur un seul champ ventral. Les sexes sont séparés ; il y a un dimorphisme sexuel ; la fécondation est interne.
On trouve environ 250 espèces de nématomorphes, regroupées en deux ordres : les Gordiens (larves parasites d’insectes en eau douce) et les Nectonématoïdes (parasites de crustacés, marins).
1 Les Gordiens.
Chez les gordiens, le pseudo-cœlome est pratiquement rempli par du mésenchyme.
Exemple : Paragordius tolosanus.
Ce parasite possède deux hôtes intermédiaires.
Les œufs donnent des larves qui vont au fond de l’eau. Elles seront ingérées par une larve d’insecte névroptère (Sialis) qui doit être mangée par un carabe (coléoptère). Une fois dans ce carabe, la larve se métamorphose en un vers adulte, libre. Ce dernier pourra réaliser la fécondation.
2 Les Nectonématoïdes.
Les nectonématoïdes sont parasites de crustacés. Les adultes sont libres et pélagiques ; mesurent environ 20 centimètres de long. On note la présence d’une double rangée de soies sur le corps. Ces soies favorisent la nage par ondulation. Le pseudo-cœlome n’est pas comblé par du mésenchyme.
Exemple : Nectonema.
C Les Rotifères.
Les rotifères ont une taille généralement inférieure à celle des ciliés (ils dépassent rarement deux millimètres). Leur corps est non-segmenté et la partie antérieure du corps est pourvue d’un appareil rotateur cilié (caractéristique de cet embranchement).
1 L’appareil rotateur.
Cet appareil est constitué par un ou deux anneaux de cils entourant une zone centrale déprimée en entonnoir (l’aire buccale). Au fond de cette aire s’ouvre la bouche. Cet appareillocomotion et à la nutrition. Il est très développé, ou réduit, ou absent, pour des formes de rotifères semi-terrestres. sert à la
La couronne extérieure postérieure est appelée paratroque (ou cingulum). La couronne interne antérieure est appelée prototroque ou trochus.
Sur la face dorsale, on peut trouver des tentacules garnis de soies sensorielles tactiles (le nombre de tentacules est fonction de l’espèce).
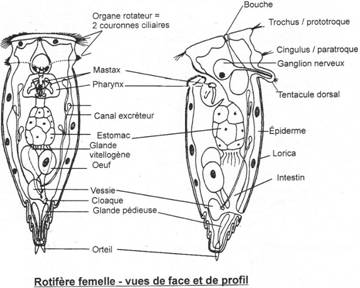
2 Le pied.
La région postérieure est appelée « le pied ». Celle-ci est terminée par deux orteils plus ou moins longs entre lesquels débouchent les glandes pédieuses qui sécrètent un liquide visqueux.
Chez certaines espèces, les orteils forment des ventouses ou des disques adhésifs qui servent aux déplacements ou à la fixation de l’animal un substrat dur.
Chez les espèces pélagiques, le pied peut être réduit ou absent. Il est très développé chez les formes primitives qui sont rampantes.
3 Paroi et musculature.
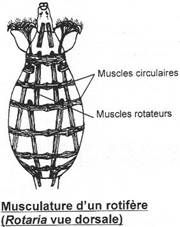
La paroi des rotifères est constituée par une cuticule (épiderme simple avec quelques faisceaux musculaires). La couche cuticulaire peut être très épaisse et divisée en plaques : on parle alors de « cuirasse » ou de lorica.
L’épiderme du pied (glandes pédieuses) sécrète une substance pour la synthèse de tubes chez les rotifères fixés. Cet épiderme donne chez les formes pélagiques, une substance gélatineuse.
La musculature est essentiellement constituée de faisceaux circulaires, longitudinaux et discontinus. Il existe des muscles rétracteurs du pied et de l’appareil rotateur.
Il existe un organe rétro-cérébral dans la région antérieure (au niveau du système nerveux central) qui a la forme d’un sac ouvert à l’extérieur. La fonction de cet organe est inconnue.
4 L’appareil digestif.
La bouche s’ouvre au fond de l’aire buccale (antérieure). Celle-ci communique avec un pharynx de deux manières :
- Directement chez les espèces prédatrices.
- Par l’intermédiaire d’un tube buccale à paroi interne ciliée.
Le pharynx est une chambre musculaire possédant un appareil masticateur (le mastax) qui est caractéristique de l’embranchement. Le mastax est formé de pièces mobiles d’origine cuticulaire. Les mâchoires sont appelées « tropi ».
On peut distinguer sept pièces cuticulaires principales : le fulcrum, les ramus (´2), les uncus (´2), les manubrium (´2).
Les ramus et le fulcrum constituent la mâchoire inférieure alors que les uncus et les manubriums constituent la mâchoire supérieure. Chez les espèces prédatrices, cet appareil peut faire saillie par l’orifice buccal pour permettre la capture et la dilacération des proies. Les pièces de cet appareil ont une morphologie différente, comme la formation des pinces ou des pièces perforantes.
Au niveau du pharynx, on trouve des glandes salivaires (entre deux et sept en fonction des espèces). L’œsophage est court et cilié. L’estomac est glandulaire ; on y trouve deux glandes gastriques. L’intestin est court, suivi d’un rectum qui débouche dans un cloaque. L’orifice cloacal est dorsal, au niveau de la descente du pied.
5 L’appareil excréteur.
L’appareil excréteur est formé par deux protonéphridies. Les canaux en partant débouchent dans une vessie contractile (ou urocyste). Elle débouche ventralement dans le cloaque. Chez certaines espèces, la vessie est absente : ce sont alors les canaux qui débouchent dans le cloaque. Cet appareil contrôle la pression interne et joue donc un rôle osmo-régulateur.
6 Système nerveux et organes des sens.
Le système nerveux est composé par un volumineux ganglion cérébral. De ce ganglion partent deux nerfs ventraux et deux terminaisons nerveuses secondaires vont innerver l’appareil rotateur, les yeux, la musculature et partent vers le corps.
Les organes sensoriels se situent au niveau de la double couronne ciliaire de l’appareil rotateur. On distingue trois types d’organes : les chémorécepteurs, les tangorécepteurs (rôle tactile), les photorécepteurs.
Les organes sensoriels sont : des soies, des papilles, des fossettes ciliées. On trouve aussi deux yeux qui vont fusionner en un œil unique rouge vif.
7 L’appareil reproducteur.
Les sexes sont séparés (gonochorisme) et le dimorphisme sexuel est très marqué (les mâles sont plus petits que les femelles). Pour de nombreuses espèces, la forme mâle n’a jamais été découverte.
L’appareil mâle :
Cet appareil est constitué d’un testicule (donc impair) poursuivit par un canal déférent qui s’ouvre au niveau du pied. Il peut y avoir formation d’une papille à l’ouverture, représentant un pénis.
L’appareil femelle :
Cet appareil est formé d’un ovaire en position ventrale, suivie d’un oviducte qui débouche dans le cloaque. L’ovaire est composé de deux parties : la partie antérieure comprend les ovules alors que la partie postérieure à un rôle vitellogène. On observe donc une séparation des fonctions gamétogènes et vitellogènes.
8 Systématique.
Les Rotifères sont divisés en trois ordres :
- L’ordre des Seisonidés.
Dans ce groupe, on trouve quelques espèces marines dont certaines qui sont épizoïquesreproduction est bisexuée, sans parthénogenèse. (vivant fixées sur d’autres organismes). Leur
- L’ordre des Bdelloïdés.
Les bdelloïdés colonisent les mousses, les lichens d’eau douce. Ils sont nageurs ou rampants. On ne connaît pas les formes mâles et il y a parthénogenèse thélytoque (femelle à femelle), indéfinie et obligatoire.
- L’ordre des Monogonontes.
Les monogonontes vivent dans les eaux douces, saumâtres, dans les mousses, les lichens. Ils peuvent être nageurs ou fixés.
9 Mode de vie.
Actuellement, on connaît environ 2000 espèces. On les trouve dans les différents milieux de vie mais particulièrement en eau douce et dans les mousses et lichens. On note quand même la présence de quelques formes parasites de végétaux et d’animaux.
Les formes aquatiques sont surtout benthiques et se déplacent à la recherche de nourriture. Les autres formes sont fixées (généralement sur des algues). Les formes planctoniques ont une morphologie externe modifiée par les conditions du milieu.
Pour une même espèce, on va avoir une morphologie différente selon le milieu de vie, ce qui entraîne des difficultés de détermination.
Les rotifères mangent des animaux, des végétaux (souvent de petite taille) qui seront attirés par la (les) couronne(s) ciliaire(s). Le mastax, chez les formes prédatrices est la première manifestation de la fragmentation des aliments dans le tube digestif.
10 Reproduction.
Les œufs ont une segmentation totale et égale.
Chez les Seisonidés, la reproduction est simple, normale. Il n’y a jamais de parthénogenèse. Chez les Bdelloïdés, toutes les femelles donnent des œufs qui se développent seulement par parthénogenèse. En revanche, les Monogonontes ont une reproduction sexuée compliquée (avec trois sortes d’œufs). Il y a alternance de générations parthénogénétiques et d’une génération sexuée.
Les femelles parthénogénétiques sont appelées « femelles amictiques ». Elles sont diploïdes et ne peuvent s’accoupler. Elles donnent des embryons à développement immédiat ne subissant pas la réduction chromatique. Ces embryons donnent des femelles diploïdes. Sous l’influence de certaines conditions du milieu (photopériode, température, pH, densité de population) apparaissent des femelles fécondables (« femelles mictiques »). Ces dernières vont donner des ovules qui émettent deux globules polaires. Si les ovules sont non fécondés, on aura des mâles haploïdes. Ces mâles pourront s’accoupler avec les femelles mictiques. La fécondation aboutit à des œufs à coquille épaisse à « œufs de durée ». Ces œufs peuvent supporter l’hiver. Quand ils se trouvent dans de bonnes conditions, ils se développent et produisent des femelles amictiques.
Les rotifères peuvent être dits monocycliques quand ils présentent plusieurs générations parthénogénétiques pendant la même saison et une génération sexuée à l’automne. Les rotifères polycycliques ont au moins deux périodes de reproduction sexuée (automne et printemps).
Pour tous, à la belle saison, il y a succession de femelles amictiques (diploïdes).
Il existe des affinités nématodiennes : symétrie bilatérale, pseudo-cœlome, cuticule, absence d’appareil respiratoire et circulaire.
Il existe aussi des différences fondamentales avec les mêmes nématodes : les nématodes ont une croissance discontinue par mues, ils n’ont pas de formations ciliaires ni d’appareil excréteur de type protonéphrien.
è Nématodes et rotifères n’ont donc pas d’ancêtre commun.
Par contre, on trouve des ressemblances remarquables avec les plathelminthes. Le système excréteur est proche, il y a séparation des fonctions gamétogènes et vitellogènes. On trouve les mêmes modalités de développement des rotifères et des turbellariés. Il y a la présence d’une ciliature assurant la locomotion. On peut aussi comparer les pièces du mastax et les pièces du pharynx de certains turbellariés.
Une parenté entre turbellariés et rotifères est donc probable.
Chapitre 4-1 :
Les Métazoaires Triploblastiques Cœlomates.
L’embranchement des annélides.
Les annélides sont des protostomiens, hyponeuriens à symétrie bilatérale. Leur corps est constitué de métamères (unités anatomiques) disposés les uns à la suite des autres. Tous les métamères de la région moyenne sont à peu près identiques : on parle alors de métamérie homonome.
Le segment antérieur est le prostomium et le segment postérieur est le pygidium : ils ne sont pas métamérisés car ils n’ont pas de vésicule cœlomique.
La chaîne ganglionnaire est ventrale, traverse tous les métamères de l’avant vers l’arrière et dans chaque métamère, on trouve une paire de ganglions nerveux.
L’appareil excréteur est métamérisé, constitué d’une paire de néphridies par segment.
L’appareil circulatoire est clos.
Le tube digestif est complet et bien différencié.
Le développement des œufs se fait par segmentation inégale, spiralée. Le développement embryonnaire donne, après la gastrulation, une larve nageuse (larve trocophore)
I La classe des Polychètes.
Les polychètes portent sur chaque segment, des parapodes (extensions latérales locomotrices) où sont implantées des soies chitineuses. Ils sont presque tous marins et les sexes sont séparés (gonochorisme).
A Morphologie externe.
La taille varie de quelques centimètres jusqu’à trois mètres. Le corps comprend trois parties :
- La région céphalique (ou prostomium) :
Cette région porte des appendices sensoriels comme les yeux (au nombre de deux) rudimentaires et sessiles. Ils portent aussi des antennes, des palpes (une paire). Les antennes et palpes assurent les fonctions tactiles et gustatives.
- Le soma :
Le soma comprend un grand nombre de segments identiques. Il commence par un métamère particulier : le péristomium. Ce dernier provient des la fusion des deux premiers métamères. Il entoure la bouche qui s’ouvre légèrement sur la face ventrale. On y trouve quatre cirres tentaculaires sur la face dorsale (à gauche et à droite). L’innervation de ces tentacules et des cirres est assurée par un ganglion de la chaîne ventrale.
- La région caudale ou pygidium :
C’est en avant du pygidium que les nouveaux métamères se forment. Le pygidium porte un anus en position terminale et deux grands cirres.
Les parapodes sont constitués de deux rames :
- Le notopode (rame dorsale) comprend un mamelon sétigère qui porte les soies. Ce dernier est encadré par deux languettes parapodiales (une dorsale inférieure et une dorsale supérieure). Dorsalement, on trouve un cirre dorsal.
- Le neuropode (rame ventrale). Cette rame est constituée d’un mamelon sétigère plus développé que le précédent : on a ainsi une languette parapodiale ventrale. Le neuropode va porter un cirre ventral.
Les rames sont soutenues par un acicule.
B Cœlome, métamérie et tégument.
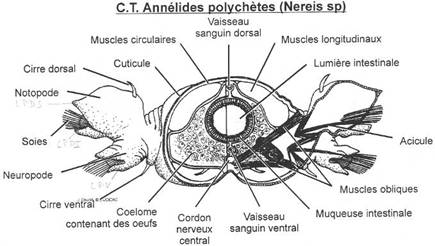
Chaque métamère est constitué de deux vésicules cœlomiques. Celles-ci sont en contactpar leurs deux feuillets splanchnopleuraux. Cet accolement donne le mésentère. Les parois cœlomiques de deux métamères consécutifs sont accolées et forment un dissépiment. La paroi du corps est formée d’une cuticule constituée de deux couches cutinisées. Elles sont striées et perpendiculaires. dans le plan médian
Sous la cuticule, on trouve un épiderme monostratifié et une membrane basalecellules à mucus utilisées chez les espèces tubicoles pour synthétiser un tube membraneux ou calcaire. d’importance variable. Cet épiderme renferme des
C L’appareil digestif.
L’appareil digestif commence par une bouche qui porte intérieurement une trompe dévaginable qui peut être inerme ou armée chez les espèces prédatrices. Dans ce dernier cas, on aura apparition de mâchoires à l’extrémité de la trompe et de paragnathes sur les cotés.
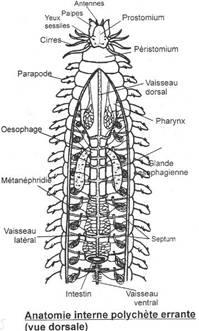
Les mâchoires servent à la capture. Une fois la proie attrapée, la trompe s’invagine et les paragnathes vont triturer cette proie.
Cet appareil se continue par une cavité buccale avec des glandes salivaires, puis un œsophage, un intestin (rectiligne) qui débouche à l’extérieur par un anus au niveau du pygidium.
D L’appareil circulatoire.
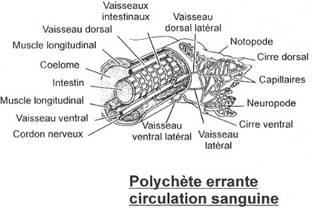
L’appareil circulatoire est clos, composé de deux vaisseaux médians (un dorsal et un ventral) réunis dans chaque segment par une paire d’anses vasculaires. Dans le tronc dorsal, le sang circule d’arrière en avant et c’est l’inverse pour le tronc ventral. Chez beaucoup de polychètes, le sang est coloré en rouge par des substances oxydables comme la chlorocruorine et/ou l’hémoglobine.
E L’appareil respiratoire.
La respiration est surtout cutanée et se fait particulièrement au niveau des parapodes. Chez certaines formes, il existe des branchies (exemple : l’arénicole) ou des extensions en panache rencontrées chez les sabelles et les serpules.
F L’appareil excréteur.
La communication entre le cœlome et le milieu extérieur se fait par les néphridies, au nombre d’une paire par segment. Chez les larves, les néphridies sont absentes, précédées par des protonéphridies. Chez les adultes, les néphridies comprennent un pavillon ciliénéphrostome). Du pavillon part un petit canal glandulaire qui traverse le dissépiment : chaque néphridie chevauche deux segments consécutifs. Ce canal glandulaire s’élargie pour donner une vessie qui débouche à l’extérieur (au niveau du segment n+1) par le néphridiopore. ouvert dans la cavité cœlomique (=
Chez de nombreuses espèces, le néphrostome ne forme pas de pavillon et est donc fermé. Quand ce pavillon est absent, on parle alors de néphridium. D’autres organes participent à l’excrétion comme la région postérieure de l’intestin et le tissu chloragogène.
G Système nerveux et organes des sens.
Chez les polychètes errantes, on trouve de nombreux organes :
- Tactiles : situés au niveau des tentacules et des palpes céphaliques.
- Nucaux : situés à la base du prostomium, chargés d’apprécier la qualité de la nourriture.
- Yeux : situés au niveau de la région céphalique ou dans d’autres régions du corps. Ces yeux sont plus évolués que ceux des planaires. Le tégument à ce niveau est transparent, permettant ainsi l’entrée de lumière.
Le système nerveux est composé d’une succession de ganglions. Les premiers ganglions (cérébroïdes) sont soudés et forme une masse unique. Ils vont innerver les organes sensoriels céphaliques (palpes, tentacules, yeux et organes nucaux). Les ganglions sont sur la face dorsale. Il se forme à l’arrière des ganglions céphaliques un collier péri-œsophagien à la suite duquel on trouve le ganglion sous œsophagien, situé dans le péristomium, sur la face ventrale. Ce dernier vient de la soudure des ganglions nerveux des deux premiers métamères. Il en part la chaîne nerveuse, constituée de deux cordons nerveux longitudinaux. Sur ces deux cordons, on trouve, dans chaque métamère, une paire de ganglions réunis par de petites commissures.
Chez les formes primitives, la chaîne nerveuse centrale est constituée de quatre cordons : on parle alors de disposition tétraneure.
H L’appareil reproducteur.
Les polychètes sont, pour la plupart, gonochoriques. Les organes génitaux ne sont pas bien différenciés. Les gonades sont de simples différenciations du revêtement péritonéal du cœlome (la splanchnopleure). Les produits génitaux tombent dans le liquide cœlomique. Chez le mâle, on obtient des spermatogonies et des ovogonies chez la femelle. Chez les mâles, les spermatogonies se multiplient et forment une masse glandulaire. Ces dernières seront libérées à l’extérieur par des canaux cœlomoductes. Quand ces cœlomoductes sont absents, les spermatogonies utilisent les canaux néphridiens ou bien, sont libérées par rupture du tégument.
I Reproduction et développement.
La fécondation est externe. La majorité des polychètes est ovipare mais il existe quelques formes vivipares. La maturité sexuelle s’accompagne de modifications morphologiques et comportementales : c’est la métamorphose ou épitoquie. Cette métamorphose entraîne des modifications surtout dans les segments postérieurs où sont stockés les gamètes. Les parapodes de ces segments développent des dilatations lamellaires. Les soies vont se développer. Dans la région antérieure, il y a aussi des modifications comme une hypertrophie des yeux et une régression des palpes et des antennes. L’épitoquie s’accompagne aussi de modifications comportementales : il y a un changement complet du milieu de vie. Par exemple, la Néréis, benthique, devient pélagique et l’on va parler d’Hétéronéréis.
Les mâles et les femelles nagent, remontent à la surface et libèrent les produits génitauxl’essaimage). L’essaimage s’effectue à des périodes bien déterminées (avec les phases lunaires). Après la reproduction, les mâles meurent. Les œufs vont se segmenter, donner une gastrula, puis, une larve trocophore ciliée, en forme de toupie, nageuse et se nourrissant de petites proies. (c’est
L’appareil digestif est différent : on a une bouche, un estomac et un intestin qui s’ouvre à l’extérieur, au niveau de l’anus. Le mésoderme est indifférencié ; au niveau de l’intestin, il va donner le mésoblaste par bourgeonnement ; les îlots mésodermiques se creusent en vésicules cœlomiques.
Chez les polychètes, il peut aussi exister une multiplication asexuée qui est présente sous diverses modalités :
- Gemmiparité : il y a formation de bourgeons qui, chacun, donneront un individu adulte. Le bourgeonnement peut se situer à différents endroits du corps d’un adulte.
- Scissiparité : des fragments d’un adulte pourront régénérer les parties manquantes. Exemple, Dodecaceria : chaque métamère peut donner un animal entier.
- Schizogamie : elle est observée chez les polychètes libérant la partie postérieure du corps. La partie antérieure va alors bourgeonner et régénérer un individu entier.
J Ecologie et systématique.
Chez les polychètes, il existe des formes libres, ectoparasites, endoparasites ou bien commensales (vivant associées avec d’autres espèces, mais pas parasites).
· Ectoparasites : ces formes parasitent les branchies de homards.
· Endoparasites : ces formes parasitent d’autres polychètes.
· Commensales : ces formes s’associent avec des annélides ou bien des éponges.
· Libres : ces formes sont marines ou d’eau saumâtre. Il existe quelques formes d’eau douce (dulçaquicoles).
On va ensuite diviser les polychètes en formes errantes et sédentaires :
Le groupe des polychètes errantes : 17 familles.
Dans ce groupe, on trouve les Néréidés. Elle est caractérisée par un grand prostomium, quatre yeux, deux antennes (ou tentacules) et deux palpes. On y trouve une trompe dévaginable avec deux mandibules et de petites dents cornées.
L’exemple des Aphroditidés : ils sont caractérisés par un corps large et court. Sur la face dorsale se différencient des cirres en forme de lame aplatie, « les élytres ».
Le groupe des polychètes sédentaires :
Ils ont deux régions au niveau du métasoma (région moyenne) : un thorax et un abdomen. La tête est souvent petite et donc peu différente du reste du corps. Les parapodes de la région moyenne sont dépourvus d’acicules. Sur ces parapodes, on a des soies en crochet au niveau des rames ventrales. On trouve aussi des branchies réparties selon une région déterminée (selon l’espèce). La trompe est sans mâchoire. Ils vivent le plus souvent dans des tubes (espèces tubicoles) et ce groupe comprend 21 familles.
Exemple des Arénicolidés. Ils ont des parapodes biramés ; les branchies sont situées sur les segments abdominaux des parapodes (exemple : Arenicola marina qui vit dans le sable, dans un tube en « U »).
Exemple des Sabellidés. Cette famille présente de longs filaments branchiaux formant une couronne tentaculaire qui peut être soit circulaire, soit en spirale. Ils vivent dans des tubes membraneux, souples et implantés verticalement dans le sable.
Exemple des Serpulidés. Ils vivent dans des tubes calcaires souvent spiralés. Ces tubes sont généralement fixés sur des coquilles de bivalves.
II La classe de clitellates.
Le groupe des clitellates comprend les oligochètes et les achètes. Les espèces de cette classe n’ont plus de parapodes. Ils présentent un renflement glandulaire se développant en période de reproduction, dans le tiers antérieur du corps : c’est le clitellum. Ce dernier se situe sur la face dorsale. Chez les lombrics, le clitellum occupe du 33ème au 38ème anneau (qui sont dans ce cas de vrais métamères). Les clitellates sont des hermaphrodites à développement direct s’effectuant dans un cocon, sans larve trocophore.
A Les Oligochètes.
1 Morphologie.
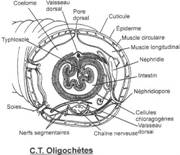
La taille des oligochètes est variable : de quelques centimètres à 3 mètres dans certains cas. Par exemple, le lombric est généralement constitué d’une centaine de métamères. L’annélation externe correspond à la métamérie. On y retrouve les différentes parties du corps des polychètes (prostomium, péristomium, soma, pygidium). Contrairement aux polychètes, les soies sont insérées dans le tégument. Chaque métamère porte quatre faisceaux de soies.
2 Le tégument.
Le tégument des oligochètes comprend un épiderme simple, constitué de trois catégories cellulaires : des cellules épithéliales, des cellules muqueuses (mucocytes), des cellules sensorielles. L’épiderme est recouvert d’une fine couche cuticulaire qui donne des reflets irisés au lombric.
Il y a modification du tégument au moment de la reproduction. Ce dernier présente un renflement glandulaire du à la multiplication des cellules muqueuses de l’épiderme.
Sous cet épiderme, on trouve deux couches de fibres musculaires superposées (une circulaire et une longitudinale). Ces couches de fibres musculaires sont réparties en quatre champs : un dorsal, un ventral et deux latéro-ventraux.
3 Le cœlome.
Le liquide cœlomique, blanchâtre, contient des cœlomocytes à fonction phagocytaireou à fonction nutritionnelle. Le cœlome communique avec la surface corporelle par l’intermédiaire de pores cœlomiques médio-dorsaux. (défense contre les bactéries)
Au niveau du tissu péritonéal, situé autour du tube digestif, on trouve des cellules de grande taille constituant le tissu chloragogène. Ces cellules sont chargées du métabolisme du glycogène et vont intervenir dans l’excrétion des déchets azotés.
4 Le tube digestif.
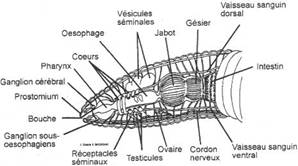
Après la bouche, on trouve une région orale située du segment 1 à 3, puis, un pharynxsegments 4 et 5). Ce pharynx comprend un renflement dorsal dévaginable (le bulbe pharyngien). Ensuite vient un œsophage dans les segments 6 à 13 qui présente des différenciations suivant le mode d’alimentation. Chez les oligochètes limicoles, la paroi interne sera ciliée. Chez les terricoles, l’épithélium est non-cilié. (
Après l’œsophage, chez les terricoles, on trouve un jabot (des segments 14 à 16) qui sert à stocker les aliments, succédé par un gésier (segments 17 à 19) permettant la trituration des aliments. Après le gésier, on a un intestin rectiligne se terminant par un anus. Cet intestin peut présenter une invagination médio-dorsale appelée typhosole. Le typhosole augmente la surface intestinale et est rempli par un tissu chloragogène.
5 L’appareil circulatoire.
L’appareil circulatoire des oligochètes est semblable à celui des polychètes (clos). On y trouve un vaisseau dorsal et un vaisseau ventral. Ces deux vaisseaux sont reliés par des anses latérales (au niveau de chaque métamère). Dans la région antérieure, les anses vont être contractiles et prendre le nom de « cœurs latéraux ». Ces « cœurs » se contractent rythmiquement mais de manière asynchrone. La contraction favorise le mouvement du flux sanguin. Chez les oligochètes primitifs, le sang est incolore. Quand on passe à des espèces un peu plus évoluées, on note la présence d’érythrocruorine (proche de l’hémoglobine).
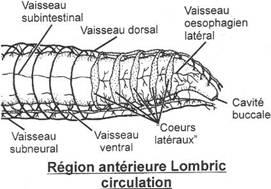
6 L’appareil respiratoire.
La plupart des oligochètes n’ont pas d’appareil respiratoire différencié. Les échanges gazeux se font alors sur toute la surface du corps, par simple diffusion pour les espèces de petite taille. Pour les espèces de grande taille, l’épiderme sera traversé de petits capillaires.
7 L’appareil excréteur.
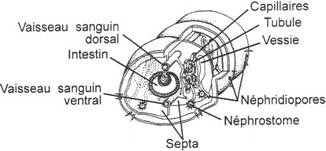
Cet appareil est semblable à celui des polychètes, avec une paire de néphridies par métamère. Les canaux sont plus longs que chez les polychètes.
8 Système nerveux et organes sensoriels.
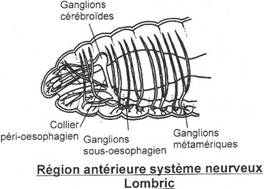
Le système nerveux rappelle en tous points celui des polychètes sauf le cerveau qui est plus simple et les organes sensoriels bien différenciés qui sont absents.
Au niveau de la chaîne nerveuse ventrale, de chaque ganglion partent trois paires de nerfs segmentaires : une paire vers la région antérieure de chaque segment et deux paires vers la partie postérieure du même segment. Les nerfs segmentaires assurent l’innervation de l’épiderme.
On note la présence de fibres géantes, multicellulaires, dont la taille peut atteindre l’équivalent de la longueur de l’animal.
9 L’appareil reproducteur.
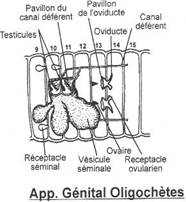
Les glandes sont nettement différenciées et localisées dans la région antérieure du corps, en avant du quinzième métamère.
L’appareil femelle comprend deux minuscules ovaires situés au niveau du dissépiment (métamères 12-13). Les ovaires lâchent leurs produits de sécrétion dans des oviductespavillons femelles, situés dans le dissépiment 13-14. L’orifice de ponte s’ouvre au niveau du quatorzième segment. captés par des
Il existe quatre réservoirs séminaux, les spermathèques, au niveau des métamères 9 et 10. Les spermathèques interviennent lors de l’accouplement et de la ponte.
L’appareil mâle est constitué de deux paires de testicules situés dans les segments 10 et 11. Sous les testicules sont présentes d’énormes vésicules séminales où les spermatogonies vont mâturer. Après la maturation, les spermatozoïdes obtenus vont utiliser deux canaux déférents s’ouvrant par des orifices mâles, au niveau de segment 15.
10 La reproduction sexuée chez les Lombrics.
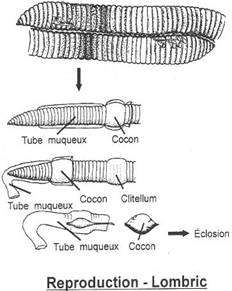
L’accouplement a lieu chez deux individus protérandriques. Ces deux animaux s’unissent tête-bêche par la face ventrale du clitellum : la région génitale de chaque individu coïncide. Les spermatozoïdes de l’un vont passer dans les réceptacles séminaux de l’autre (et inversement). L’accolement est favorisé par la sécrétion de mucus (au niveau du clitellum).
Une fois l’échange de spermatozoïdes réalisé, les deux individus vont se séparer. Chacun synthétise un cocon au niveau du clitellum où sont déposés les ovocytes. Le lombric va reculer en laissant le cocon sur place. Une fois ce cocon au niveau des orifices mâles, les spermatozoïdes y seront libérés : c’est la fécondation.
Le développement est direct : l’embryon donne un jeune ressemblant à l’adulte.
Il existe chez les oligochètes asexués, une multiplication par scissiparité (chez les limicoles), se faisant par deux phénomènes : architomie ou paratomie.
11 Biologie des Oligochètes.
La plupart des oligochètes se déplacent par reptations péristaltiques : par des contractions alternatives des muscles longitudinaux et transversaux. Les soies jouent un rôle d’accrochage. Les oligochètes ont la possibilité de passer dans des orifices de diamètre inférieur à celui de leur corps, grâce à la mobilité du liquide cœlomique.
12 Régime alimentaire.
Les Oligochètes se nourrissent de détritus organiques variés. Ils peuvent être aussi bien marins que d’eaux douces ou terrestres, dans des sols humides. Il existe quelques espèces prédatrices et quelques espèces parasites. On peut distinguer trois catégories d’oligochètes :
- Les Oligochètes Limicoles nageurs : ils vivent sur des plantes immergées.
- Les Oligochètes limicoles fouisseurs : ils vivent dans les sédiments des lacs et des rivières.
- Les Oligochètes strictement terrestres : ces formes ont un rôle important dans la formation des sols : rôle mécanique et chimique. On estime la quantité de lombrics par hectare à une tonne (par hectare). Ils peuvent ingérer 200 à 300 tonnes de terre par an. Ils produisent la plus grande quantité d’humus recouvrant la terre. Ils ont aussi un rôle d‘oxygénation et de drainage des sols.
B Les Achètes.
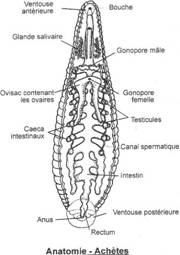
Les achètes sont majoritairement représentés par les sang-sues. Ces formes peuvent être libres ou ectoparasites. On les trouve principalement en eau douce. Il existe quand même quelques formes marines et terrestres.
1 Le cas de Hirudo medicinalis.
Cette espèce ne possède ni parapode ni soie. Le nombre de segments métamérique est fixe (33), auquel on rajoute le prostomium. Les anneaux externes ne correspondent pas à la métamérie interne : on observe 102 anneaux externes chez la sang-sue.
La région antérieure porte les yeux, rudimentaires (cinq paires), sur le premier anneau des cinq premiers métamères. Les métamères 1 et 2 n’ont qu’un anneau externe. Le troisième en possède deux. Les 4 et 5 en ont trois chacun. On trouve aussi une ventouse buccale en arrière du prostomium et une ventouse à l’extrémité postérieure du corps qui est non perforée.
On trouve un clitellum moins marqué que chez les oligochètes. Ce clitellum est situé du neuvième au douzième métamère. Là, s’ouvrent les orifices génitaux males et femelles en position médio-ventrale. Les orifices males sont sur le métamère 10 (M10) et les orifices femelles sont sur M11.
2 Le tégument.
L’épiderme est mono-stratifié, recouvert par une cuticule plus ou moins épaisse, selon la région du corps et selon l’espèce. La cuticule s’interrompt au niveau d’un petit orifice correspondant aux glandes cutanées.
Le derme est formé d’un tissu conjonctif. Sous le derme, on trouve une musculature très développée, présente sous trois formes : circulaire, longitudinale et dorso-ventrale.
Le cœlome est réduit car un tissu de remplissage (tissu botryoïdal) est présent : il a le même rôle que le tissu chloragogène des oligochètes.
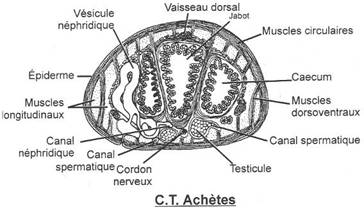
3 L’appareil digestif.
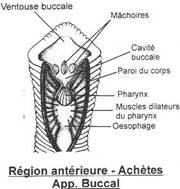
L’appareil digestif est composé de :
- La partie antérieure, d’origine ectodermique, avec une cavité buccale, le pharynx et l’œsophage.
- La partie moyenne correspond à l’estomac, d’origine endodermique. On y trouve des cæcums digestifs dont la taille croit de M7 à M18.
- La partie postérieure, d’origine endodermique, est la partie où se déroule la digestion. Elle est terminée par le rectum (ou canal rectal), débouchant à l’extérieur par un anus en position dorsale, dans M26.
4 L’appareil circulatoire.
Quand l’appareil circulatoire est présent, il forme un système vasculaire clos, à parois contractiles. Il est constitué d’un vaisseau dorsal dans lequel le sang circule d’arrière en avant, et d’un vaisseau ventral. Les deux vaisseaux sont réunis aux extrémités par de nombreuses anses vasculaires.
Certains achètes n’ont pas d’appareil circulatoire ; c’est alors le système cœlomique et la lymphe qui vont jouer le rôle du sang.
5 L’appareil respiratoire.
Il n’existe pas d’appareil respiratoire. Les échanges gazeux se font au travers de l’épiderme. Certaines espèces parasites de poissons marins ont des replis cutanés fonctionnant comme des branchies.
6 L’appareil excréteur.
L’appareil excréteur comporte une quinzaine de paires de métanéphridies qui s’ouvrent à l’extérieur au niveau de néphridiopores, situés ventralement, au niveau de M7 à M23.
7 Le système nerveux et organes sensoriels.
Le système nerveux est plus complexe que chez les polychètes et les oligochètes. Il est constitué de 34 paires de ganglions répartis ainsi : les six premières paires fusionnent pour donner le « cerveau » et une masse nerveuse sous-œsophagienne. Les 21 paires de ganglions suivants sont réparties le long de la chaîne nerveuse. Les sept dernières paires de ganglions fusionnent en une masse nerveuse pygidiale.
Les organes sensoriels sont relativement réduits. On trouve des yeux céphaliques, un grand nombre de papilles sensitives et des cellules tactiles.
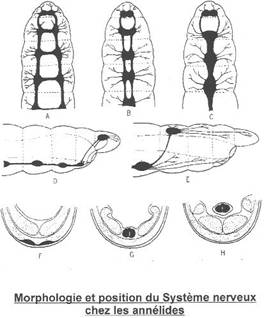
De A à C, on observe une fusion des ganglions puis des cordons nerveux.
En D et E, on voit les ganglions cérébroïdes qui reculent en arrière du prostomium, vers M3. Les ganglions sous-œsophagiens sont repoussés dans M4. On a aussi un développement des prolongements des chaînes nerveuses vers l’avant pour les organes sensoriels et les muscles.
De F à H, on observe un enfoncement de la chaîne nerveuse à l’intérieur de l’animal. Primitivement (en F), la chaîne nerveuse est sous-épidermique ; elle s’enfonce ensuite (en G) dans les muscles longitudinaux (sous-dermiques) et finie (en H) dans la chaîne de la cavité cœlomique.
8 Les appareils génitaux.
· L’appareil male.
L’appareil male se compose d’un nombre variable de testicules selon les espèces. Chez Hirudo, on trouve dix paires de testicules de M12 à M21. Ces testicules sont reliés par de longs spermiductes longitudinaux. Ils s’élargissent en vésicules séminales pour se terminer en canal éjaculateur (un seul orifice male).
· L’appareil femelle.
L’appareil femelle est constitué d’une paire d’ovaires qui sont contenus dans des ovisacs. C’est là que les cellules germinales se forment. De ces ovaires partent deux oviductes courts, débouchant dans deux utérus. Ces deux derniers confluent dans un vagin qui s’ouvre à l’extérieur par un pore génital unique (M11).
9 Les différents ordres d’Achètes.
L’ordre des Acanthobdelliformes.
Dans cet ordre, on ne trouve qu’une seule espèce : Acanthobdella peledina. Elle est parasite de poissons salmonidés (truites, saumons) du lac Baïkal. Cette espèce possède des caractères communs avec les oligochètes, comme la présence de soies sur le corps et l’absence de ventouse antérieure.
L’ordre des Rhyncobdelliformes.
Cet ordre regroupe les sang-sues à trompe dévaginable. Toutes les formes sont aquatiques. On peut distinguer deux familles. La famille des Glossiphoniidés comprend des espèces, toutes parasites de mollusques ou d’amphibiens. La seconde famille, celle des Pissicolidés comprend des individus parasites de poissons.
L’ordre des Gnathobdelliformes.
Cet ordre regroupe des sang-sues aquatiques ou terrestres présentant trois mâchoires dentées au niveau du pharynx. On y trouve Hirudo officinalis.
L’ordre des Pharyngobdelliformes.
Ces individus sont d’eaux douces ou terrestres de milieux humides. Ils sont presque tous prédateurs. Leur pharynx est long, sans mâchoire.
10 Biologie et Ecologie.
Il n’existe pas de multiplication asexuée chez les achètes. La fécondation sexuelle est soitréciproque, soit unilatérale (= autofécondation). Les œufs fécondés sont pondus dans un cocon selon la même modalité que chez les oligochètes. Le cocon contient un liquide « albumineux » servant de nourriture à l’embryon.
· Régime alimentaire.
Il existe de nombreuses formes parasites ou prédatrices. Pour les formes parasites, on parle de parasitisme temporaire car ces sang-sues se détachent de l’hôte, une fois le repas terminé. Elles sont encore utilisées en chirurgie réparatrice pour relancer la circulation veineuse dans les membres accidentés (elles créent un appel du sang).
III les Pogonophores.
L’embranchement des annélides comprend en plus un autre groupe, les pogonophores, uniquement constitué d’espèces (peu nombreuses) cantonnées aux fonds abyssaux.
A Morphologie externe.
Les pogonophores ont souvent une forme de fil fin (inférieur à 1mm) d’une longueur d’environ 8cm. Certaines espèces peuvent mesurer jusqu’à 2,5mm de diamètre pour 30cm de long. On a même trouvé un individu de 1,5m de long et de 5cm de diamètre.
Ces animaux vivent dans des tubes et possèdent des couleurs vives et contrastées. La partie antérieure porte de nombreux tentacules garnis de pinnules. Cette partie avant est appelée le « prosome ».
La partie moyenne (le mésosome) et la partie terminale (le métasome) portent des papilles et des plaquettes chitineuses que l’animal utilise pour se déplacer dans son tube.
B Morphologie interne.
Le système nerveux est cutané ; il n’y a pas d’organes des sens ; le système circulatoire est clos et formé de deux vaisseaux (un dorsal et un ventral). Dans le vaisseau dorsal, le sang circule d’avant en arrière.
Ces animaux sont gonochoriques. La fécondation se fait dans le tube de la femelle. Les œufs se développeront dans la partie antérieure du tube.
Les pogonophores sont caractérisés par l’absence de tube digestif bien que non parasites. Deux hypothèses existent pour expliquer le mode d’alimentation :
- Il y aurait absorption, au niveau des cellules épithéliales du tégument, de substances dégradées par les bactéries qui vivent en contact avec eux.
- Il y aurait une digestion externe de particules qui seraient ensuite récoltées par les tentacules pinées. Toutefois, aucune glande enzymatique pour la digestion externe n’a pu être mise en évidence.
Chapitre 4-2 :
Les Métazoaires Triploblastiques Cœlomates.
L’embranchement des Mollusques.
Cet embranchement compte plus de 130 000 espèces et occupe donc une place importante dans le règne animal par le nombre (d’espèces) et par le rôle écologique. Certaines espèces servent de nourriture pour l’Homme. Ce sont les seuls invertébrés avec les crustacés à être consommés (très fréquemment).
On distingue sept classes de mollusques : les Monoplacophores, les Aplacophores, les Polyplacophores, les Scaphopodes, les Lamellibranches (= Bivalves), les Gastéropodes et les Céphalopodes. Nous verrons en détail les trois dernières classes.
Chez les mollusques, on parle de radiation adaptative car, à partir d’un ancêtre commun, une grande quantité d’espèces a évolué. Les sept classes ont un ancêtre commun. Ces espèces ont principalement conquis le milieu marin où l’on trouve des mollusques dans les zones de balancement de marées, dans le milieu pélagique (en pleine eau), dans les substrats meubles (comme le sable), dans les substrats durs (les rochers) et à de grandes profondeurs marines.
Seuls deux groupes se sont adaptés avec succès à l’eau douce : les gastéropodes et les lamellibranches ; et quelques gastéropodes sont même passés à la vie terrestre. On pense que ces mollusques ont été les premiers à sortir de l’eau.
Bien que les espèces de ces diverses classes soient différentes, on peut quand même définir des caractères communs à ces sept groupes.
I Les caractères communs.
- Il n’y a jamais de segmentation, sauf chez les monoplacophores.
- Les mollusques sont tous fortement céphalisés avec la présence d’une tête et d’organes sensoriels spécialisés.
- Le cœlome est réduit et la paroi du corps est épaisse et musculeuse.
- La partie ventrale de la paroi du corps est transformée en un pied musculeux habituellement utilisé pour la locomotion.
- La partie dorsale du corps va former le manteau (ou pallium). Celui-ci sécrète une coquille et délimite la cavité palléale dans laquelle sont situées les branchies.
- Dans la cavité palléale s’ouvrent l’anus et tous les autres pores excréteurs.
- Le tube digestif est bien développé et régionalisé. La cavité buccale contient presque toujours un organe tubulaire d’alimentation (= la radula). Cette dernière est armée de dents et fonctionne comme une râpe. On trouve ensuite un œsophage qui est composé de régions spécialisées : une région pour le stockage de la nourriture et une région pour le morcellement de celle-ci. Il vient ensuite un estomac associé à une paire de glandes digestives. La partie postérieure de ce tube digestif forme l’intestin, souvent long et terminé par un anus.
- On trouve presque toujours un système circulatoire avec un cœur bien défini. Ce cœur est constitué d’une ou plusieurs oreillettes et d’un ventricule médian. Le sang circule dans les vaisseaux (sanguins) mais aussi dans les espaces inter-viscéraux entourant le cœlome. Le pigment respiratoire est l’hémocyanine.
- On trouve une paire de reins pouvant fusionner. Ils sont en liaison étroite avec la cavité péricardique.
- Le système nerveux. On trouve un anneau nerveux circum-entérique (= péri œsophagien). A cet anneau sont associées deux paires de connectifs (nerveux). Une des deux paires innerve le pied (connectifs cérébropédieux), l’autre innerve les viscères et le manteau (connectifs cérébropleuraux). Sur l’anneau nerveux et les connectifs, on trouve des ganglions (nerveux) plus de vastes réseaux nerveux sous-épidermiques.
- Les œufs sont généralement de taille réduite, avec peu de vitellus. La segmentation est spirale. La larve caractéristique des mollusques est une larve trocophore (pour les mollusques primitifs) ou une larve velligère (pour les mollusques moins primitifs). Elles sont toutes les deux très proches.
Tous ces caractères ne sont pas forcément présents en même temps dans les sept classes.
A Le mollusque primitif.
Ce mollusque a une tête bien définie, avec des tentacules céphaliques portant des yeux. La face ventrale est modifiée en sole de reptation : le pied. La partie dorsale (ou masse viscérale) est recouverte par un repli de la paroi du corps (le manteau) qui peut sécréter soit une coquille soit des spicules calcaires.
Vers l’extrémité postérieure, le manteau délimite une cavité (palléale) dans laquelle on trouve les branchies (ou cténidies) et l’ouverture de l’anus et des néphridiopores. La structure branchiale est avec un axe central, à partir duquel partent, de chaque côté, des filaments aplatis.
Sous la tête, on trouve l’ouverture de la bouche avec la cavité buccale, musculeuse. A ce niveau, on trouve des glandes salivaires qui produisent un mucus servant à agglutiner les fragments de nourriture pour faciliter le passage de cette nourriture dans l’œsophage. Il s’y ouvre un sac radulaire, contenant la radula qui est un ruban garni de dents, pouvant être projeté hors de la bouche et utilisé par l’animal, pour arracher la nourriture du substrat sur lequel elle se trouve.
La paroi interne de l’œsophage est tapissée de cils qui facilitent le passage de la nourriture dans l’estomac.
Dans l’estomac, on a une région sclérotinisée, où la nourriture la plus dure peut être broyée. La cœlome comportait deux compartiments largement communicant et dont l’un entoure le cœur et l’autre, une partie de l’intestin. Dans ce cœlome, on trouve aussi les gonades. C’est aussi à ce niveau cœlomique, que sont collectés les déchets, au niveau des néphrostomes. Les déchets sont libérés dans la cavité palléale par l’intermédiaire des néphridiopores.
B Les points communs entre les sept classes actuelles.
En partant de ce mollusque primitif, en aplatissant la masse viscérale et en prolongeant la cavité palléale en forme de sillon entourant le pied, on obtient la structure des polyplacophores.
Pour avoir un lamellibranche, il faut élargir le manteau, le diviser en deux moitiés symétriques : cette transformation définie alors une cavité palléale immense. La surface des branchies est beaucoup plus importante. Il faut étirer le pied vers le bas de la région antérieure. Le pied donne une lame et va pouvoir aller à l’extérieur de l’animal en passant entre les deux moitiés du manteau.
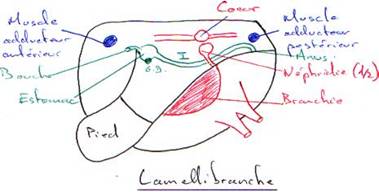
Le passage à la forme scaphopode se fait en prenant un lamellibranche, en soudant les bords inférieurs du manteau et en l’étirant en longueur (sens antéro-postérieur). On a finalement une structure tubulaire à l’intérieur de laquelle le corps va s’allonger.

Pour avoir un gastéropode ou un céphalopode, le processus est différent. La masse viscérale doit se développer en hauteur et donner une bosse. Au fur et à mesure qu’elle s’élève, elle va avoir tendance à s’enrouler. Tous les céphalopodes n’ont pas de masse viscérale enroulée ; cependant, les premières espèces de céphalopodes et la forme nautilus avaient un corps enroulé.
La différence importante entre les céphalopodes et les gastéropodes vient de la tête et du pied qui sont mieux individualisés chez les gastéropodes. Il se rapproche plus de la forme ancestrale.
C Quelques caractères communs aux sept classes.
Tous les mollusques sont des métazoaires triploblastiques cœlomates. Le cœlome, constitué au début par une paire de sacs cœlomiques, se transforme en trois cavités communiquant entre elles. La cavité génitale entoure les gonades ; la cavité péricardiquecœur ; un néphrocœle entoure les néphridies. Les deux dernières cavités peuvent fusionner pour donner une cavité réno-céphalique. entoure le
La cavité palléale est délimitée par le pallium. Elle joue un rôle important chez les mollusques. Il y débouche le tube digestif par l’intermédiaire de l’anus, les néphridiopores. Cette cavité renferme le plus souvent une paire de branchies constituées de cellules ciliées. La vibration de ces cils entraîne un fort courant d’eau, obligatoire pour la respiration et même parfois, pour l’alimentation (surtout chez les lamellibranches).
On peut aussi trouver, dans la cavité palléale, des organes sensoriels comme les osphradies, qui permettent à l’animal d’apprécier la qualité de l’eau « inhalée ». On a aussi des glandes sécrétant une substance qui permet le nettoyage de cette cavité : les glandes hypobranchiales.
Cette cavité se transforme chez les gastéropodes terrestres (les Pulmonés). Elle se renferme en un sac (le « poumon ») qui communique avec le milieu extérieur par un orifice pulmonaire : le pneumostome. On pourra observer au niveau du manteau, une ou plusieurs soudures délimitant des ouvertures avec le milieu extérieur. On peut y trouver des prolongements tubulaires : les siphons.
1 L’appareil digestif des mollusques.
Le tube digestif commence par une bouche s’ouvrant sur le bulbe buccal, au niveau duquel se trouve la radula. Celle-ci est constituée de dents chitinisées fonctionnant comme une râpe. La radula est enveloppée par une gaine constamment régénérée au fur et à mesure de la vie de l’animal.
Chez les gastéropodes, le nombre, la forme et la disposition des dents sont fonctions du régime alimentaire (les dents sont un des caractères de la systématique de la classe des gastéropodes).
Chez certains mollusques, on va trouver des mâchoires, par exemple, le bec de perroquet des céphalopodes.
Au niveau du bulbe buccal vont arriver deux glandes salivaires. La bouche se prolonge d’un œsophage puis, d’un estomac on l’on trouve l’arrivée d’une glande digestive (dans l’estomac). Il est parfois appelé « hépato-pancréas » mais ce terme est à proscrire. Il suit l’intestin qui est souvent long, débouchant dans la cavité palléale par un anus.
Un élément particulier de ce tube digestif est le protostyle qui permet la trituration et la digestion des aliments. Il se trouve dans l’intestin. Comme la radula, il se régénère au cours de son utilisation. Il a une forme de bâtonnet et a deux rôles dans la digestion :
- Un rôle chimique par l’action d’enzymes.
- Un rôle mécanique : il peut tourner rapidement sur lui-même et provoquer un écrasement.
2 La coquille.
Cf. TP.
3 Le système nerveux.
Le système nerveux est constitué de plusieurs paires de ganglions reliés entre eux par descommissures (pour les mêmes paires) ou par des connectifs nerveux (pour les ganglions de paires différentes).
La disposition des ganglions est particulière et dite « en triangle » (ganglions cérébroïdes, pleuraux et pédieux). Excentrés par rapport au triangle, on trouve les ganglions viscéraux reliés aux ganglions pleuraux.
Cette disposition est plus ou moins visible chez toutes les classes de gastéropodes (il peut y avoir fusion des ganglions et/ou liaisons entre ganglions). On pourra trouver une fusion totale des ganglions pour former un « cerveau ». Les céphalopodes sont les mollusques les plus évolués.
4 L’appareil circulatoire.
L’appareil circulatoire comporte un cœur enveloppé par un péricarde. Le cœur est constitué d’un ventricule et de deux oreillettes. Chaque oreillette est en communication avec une branchie. Le sang d’une oreillette va vers le ventricule avant d’aller irriguer les différents organes par l’intermédiaire des artères. Tous les mollusques ont un système circulatoire complètement clos. Dans certains cas, il peut y avoir disparition d’une oreillette (très souvent chez les gastéropodes). Le pigment respiratoire est l’hémocyanine. Il renferme du cuivre qui correspond à l’élément métallique de l’hème.
5 L’appareil respiratoire.
L’appareil respiratoire est étroitement lié à la cavité palléale. Il est constitué de branchiespoumon chez les gastéropodes terrestres (les pulmonés). qui vont être remplacées par un
Les branchies ont une forme variable. Elles constituent le lieu d’échanges respiratoires chez les mollusques aquatiques.
6 L’appareil excréteur.
L’appareil excréteur est représenté par deux reins qui sont à l’origine de la formation des cœlomoductes à paroi excrétrice. Ces cœlomoductes communiquent avec la cavité péricardique et s’ouvrent à l’extérieur au niveau de la cavité palléale par l’orifice excréteur.
La communication entre rein et péricarde se trouve dans la plupart des groupes. Elle est absente chez les gastéropodes terrestres.
7 L’appareil génital.
Les sexes sont généralement séparés chez les mollusques mais on trouve des exemples d’hermaphrodisme et quelques cas d’ambisexualité (inversion du sexe durant la vie).
Les gamètes, produits par les gonades, passent dans la cavité péricardique puis dans les canaux excréteurs (cœlomoductes) pour être amenés dans la cavité palléale.
La fécondation est externe pour les lamellibranches. Il y a accouplement chez les céphalopodes et les gastéropodes.
Après la fécondation, l’œuf subit une segmentation spirale. Cet œuf donnera dans la majorité des cas une larve trocophore (libre et nageuse). Il existe quelques exceptions : la larve trocophore peut être modifiée rapidement pour former la larve velligère (avec une couronne ciliée développée à prototroche). Sur la face dorsale de cette larve se présente rapidement une coquille.
Les céphalopodes n’ont pas de développement stade larvaire libre : le développement embryonnaire a lieu chez la femelle.
Les gastéropodes n’ont pas tous de larve velligère.
II Etude de classes.
Avant, les amphineures regroupaient les polyplacophores et les aplacophores. Ces deux classes sont caractérisées par l’absence de torsion du tube digestif : la bouche et l’anus sont terminaux et opposés.
Le manteau va synthétiser des plaques calcaires qui donnent la carapace ou bien des spicules. Les aplacophores comprennent des spicules dans leur masse viscérale.
A Les Aplacophores.
Les aplacophores sont de petits animaux vivant dans le sol ou vivant fixés sur des plantes ou d’autres mollusques.
B Les Polyplacophores.
Un des représentants de ce groupe est le Chiton. On le trouve sur les rochers d’Atlantique ou de Méditerranée. Le corps est aplati dorso-ventralement. La coque dorsale est constituée de huit plaques calcaires articulées entre elles. Ventralement, on trouve un pied large ou sole de reptation. La cavité palléale va former le sillon palléal autour du pied. Dans le sillon, on trouve les branchies.
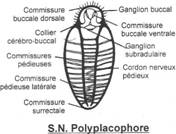
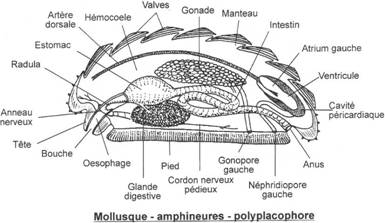
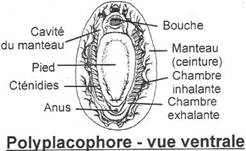
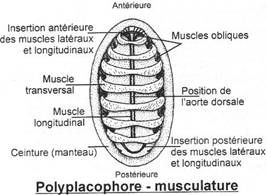
Le chiton possède une gonade qui provient de la fusion des deux gonades primitives. Il reste deux conduits génitaux. La fécondation est externe et les sexes sont séparés.
C Les Monoplacophores.
Au début des années 50, on ne les trouvait qu’à l’état de fossiles. En 1952, on a trouvé des exemplaires vivants du genre Néopilina à plus de 4000 mètres de profondeur, dans le pacifique, au niveau du Mexique.
C’est un animal circulaire d’environ quatre centimètres de diamètre, montrant une symétrie bilatérale nette. Néopilina possède une coque dorsale mince et est caractérisé par une répétition d’un certain nombre d’organes pouvant faire penser à une métamérisation de l’animal. Dans la région antérieure s’ouvre la bouche, entourée par des palpes ciliés et des tentacules buccaux. Le pied est circulaire et contient huit paires de muscles rétracteurs.
Dans le sillon palléal, on trouve cinq paires de branchies. Dans ce sillon débouchent six paires de néphridies et deux paires de gonades.
Chez Néopilina, les sexes sont séparés et la fécondation externe.
Le cœur est composé d’un ventricule et de deux oreillettes. Le cœlome est plus développé que chez les autres mollusques.
Néopilina galathea possède des caractères primitifs comme le sillon palléal. Cela le rapproche du mollusque ancestral type. Cependant, cette espèce se distingue des autres mollusques par la répétition d’un certain nombre d’organes (musculature, néphridies, …) : c’est une forme de métamérie.
è Les monoplacophores sont rapprochables des annélides. On y trouve une segmentation spirale des œufs ainsi que quelques ressemblances larvaires. Ils descendraient de formes annélidiennes métamérisées. Cette hypothèse a été abandonnée : maintenant, on pense qu’ils viendraient de plathelminthes turbellariés primitifs. De nombreux travaux de taxonomie moléculaire sont effectués pour connaître la phylogénie de cette classe.
D Les Scaphopodes.
Les scaphopodes sont de petits animaux marins. Le plus connu d’entre eux est le dentaleDentalium) qui vit dans les sables. Son corps est allongé et enveloppé d’un manteau tubulaire sécrétant une coquille en forme de cône. (
On va trouver deux ouvertures :
- Une ouverture large dans la région antérieure (dans le sable).
- Une ouverture étroite, qui fait saillie hors du sédiment.
C’est par l’ouverture antérieure que s’ouvre la bouche. Celle-ci est entourée de tentaculescaptacules ». Le pied est peu développé et a un rôle fouisseur. appelés «
La cavité palléale s’ouvre par l’orifice le plus étroit (postérieur).
Les branchies ont disparues et les échanges respiratoires se font alors au niveau du manteau.
Le système nerveux est constitué de quatre paires de ganglions (cérébroïdes, pleuraux, pédieux et viscéraux).
Les sexes sont séparés et l’on ne trouve qu’une gonade.
Ce groupe est rapproché des gastéropodes car leur coquille est unique et leur masse viscérale se développe en hauteur. Ils sont aussi rapprochés des lamellibranches car leur tête est réduite, leur pied fouisseur et la coquille, chez la larve de scaphopode, est bivalve.
E Les Gastéropodes.
La classe des gastéropodes regroupe les ¾ des espèces de mollusques. Ils se distinguent par la disparition de la symétrie bilatérale au profit d’un enroulement hélicoïdal de la masse viscérale.
1 Caractères généraux.
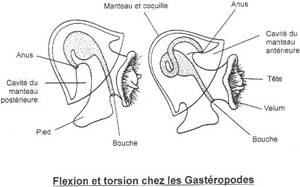
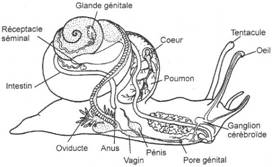
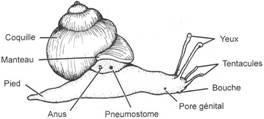
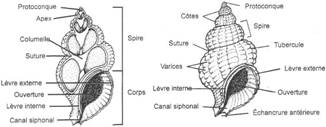
La tête est bien individualisée, porte des organes sensoriels comme les yeux ou les tentacules.
Le pied est très musculeux et souvent appelé « sole de reptation ». Celle-ci sert à l’appui de l’ensemble de la masse viscérale qui est protégée par une coquille.
La coquille est formée par un tube conique entouré en spirale autour d’un axe rigide (la columelle). Cette columelle peut être creuse ou pleine.
Au cours du développement larvaire, on observe trois phénomènes caractéristiques de cette classe : la flexion endogastrique, la torsion et la spiralisation. La flexion endogastrique va provoquer un changement d’orientation au niveau de la cavité palléale. La torsion va amener cette cavité en avant, du côté dorsal. La spiralisation va entraîner l’enroulement de toute la masse viscérale qui se retrouve dans la coquille. Elle entraîne aussi la perte de la symétrie bilatérale. Toutefois, il faut noter que tous les gastéropodes ne subissent pas la spiralisation.
C’est la torsion qui permet la différenciation des sous-groupes de gastéropodes.
2 La sous-Classe des Prosobranches.
Les Prosobranches ont subit une torsion complète (180°). La cavité palléale se retrouve en avant (elle comporte les branchies). Les sexes sont séparés ; la coquille est bien développée et peut être refermée par un opercule rigide d’origine organique ou calcaire. On y trouve 3 ordres :
- Les Archéogastéropodes. C’est le groupe le plus primitif. Les individus le composant sont tous marins, généralement herbivores. Exemples : Patelle, Haliotis, Ormeaux.
- Les Mésogastéropodes. Ce groupe est celui qui comprend le plus d’individus et c’est aussi le plus varié des Prosobranches. Ces espèces ont en général une branchie, un rein et une oreillette. Ils sont souvent herbivores mais on y trouve quand même des carnivores se nourrissant de lamellibranches (comme les bigorneaux).
- Les Néogastéropodes : Ce sont les plus évolués. Ils sont carnivores et certains possèdent un venin toxique pouvant être mortel pour l’Homme. Exemples : Buccin, Murex.
3 La sous-Classe des Opisthobranches.
La torsion des opistobranches est incomplète, ne dépassant pas 90°. Ils n’ont en général qu’un rein, qu’une oreillette, qu’une branchie. La coquille est réduite, souvent interne et parfois absente. On y distingue deux ordres :
- Les Tectibranches. Exemple : le lièvre de mer (genre Apbysia). Il y a un développement des expansions latérales du pied : les parapodies. La coquille est très mince et le manteau tend à la recouvrir. Les parapodies s’hyper-développent et fonctionnent comme des nageoires ou des ailes è aptitude à la nage. Ce lièvre de mer est un brouteur d’algue.
- Les Nudibranches. Il y a disparition totale de la coquille et de la cavité palléale. Les branchies sont remplacées par des expansions du tégument : les papilles. Ces nudibranches se nourrissent de cnidaires mais ils ne digèrent pas les cnidoblastes ; ils stockent parfois ces derniers au niveau des papilles, ce qui les protège.
4 La sous-Classe des Pulmonés.
On trouve les pulmonés en eau douce ou en milieux terrestres ; exceptionnellement en mer. Chez les formes terrestres, les branchies disparaissent. Le plafond de la cavité palléale forme un poumon. L’ouverture de la cavité se rétrécie et elle communique à l’extérieur par le pneumostome.
L’appareil génital est hermaphrodite. Il y a accouplement et fécondation croisée mais aussi auto-fécondation possible. On distingue deux ordres :
- Les Basomatophores. Ils ont les yeux situés à la base des tentacules qui ne sont pas invaginables. La coquille est bien développée. Ce sont les pulmonés qui retournent à la vie aquatique. On les trouve en eau douce. Exemple : Planorbe dont la coquille s’enroule dans un plan.
- Les Stylomatophores. Ils sont presque tous terrestres (genre Hélix). La coquille extérieure est bien développée. La columelle est creuse chez l’escargot de Bourgogne. Chez les limaces, la coquille est réduite voir interne.
F La classe des Lamellibranches (ou Bivalves).
Cette classe ne comprend que des individus marins.
1 Caractères généraux.
· Le manteau.
Le manteau est développé et donne deux lobes symétriques enveloppant la totalité du corps et constitue la cavité palléale. Les deux lobes pourront être libres sur toute leur surface sauf dorsalement où l’on trouve une soudure au niveau de la charnière. Ces deux lobes du manteau vont laisser une empreinte sur la face interne des valves : c’est la coquilleintégripalliée (empreinte continue). Chez quelques formes, il existe d’autres types d’empreintes (chez les lamellibranches fouisseurs) : une ou plusieurs soudures ventrales. Quand on a deux soudures ventrales, il y a deux orifices dont un est inhalant et l’autre exhalant. Chez les fouisseurs les mieux adaptés, les orifices sont prolongés par dessiphons. L’empreinte laissée est donc discontinue : c’est la coquille sinupalliée.
· Les muscles adducteurs.
On trouve généralement deux muscles adducteurs (un antérieur et un postérieur) : espèce dimyaire. Ces deux muscles traversent de part en part l’animal et s’insèrent sur la face interne des deux valves. La fixation laisse des empreintes.
Le rôle de ces muscles est de fermer ou de rapprocher rapidement les deux valves.
Il existe quelques formes avec un muscle adducteur postérieur unique : espèce monomyaire.
· La tête.
On note une absence de tête bien différenciée. Il n’y a donc pas d’organes sensorielscéphaliques. Ces organes se retrouvent plutôt sur les bords du manteau.
· Le tube digestif.
La partie antérieure du tube digestif ne comporte pas de mâchoire, ni de radula, ni de pharynx. La bouche est entourée par des palpes (deux paires de palpes labiaux) et s’ouvre sur l’œsophage.
· L’appareil respiratoire.
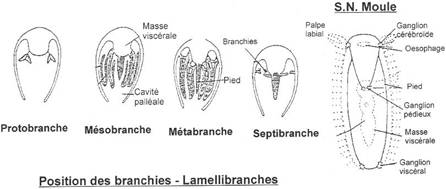
Les branchies sont recouvertes par un épithélium cilié dont les battements créent un puissant courant entrant d’eau. Il favorise la respiration par apport d’eau oxygénée et l’entrée de petites particules alimentaires qui pourront progresser jusqu’à la bouche. Le pouvoir filtrant de ces animaux est très important. Par exemple, Corbicula (1 centimètre de diamètre) filtre une dizaine de litres d’eau par jour.
· La reproduction.
Les males et les femelles émettent leurs produits génitaux dans l’eau (fécondation externe). Il existe un phénomène particulier pour les lamellibranches d’eau douce : après le développement embryonnaire, on observe le développement d’une larve glochidieparasite) qui se fixe sur les branchies ou les nageoires de poissons. Cette fixation confère aux mollusques d’eau douce, un grand pouvoir de dissémination. (
La première classification des lamellibranches est basée sur la structure et la forme des branchies.
2 L’ordre des Protobranches.
Les branchies sont constituées de filaments simples et le manteau est largement ouvert. Exemple Nucula sp.
3 L’ordre des Mésobranches (ou filibranches).
Les branchies sont plus complexes, constituées de filaments directs et réfléchis. Ils sont en liaison par une jonction simple. Les branchies prennent une forme de « W ».
Les deux muscles adducteurs sont présents mais le muscle antérieur est réduit. On trouve une soudure dorsale (la charnière). Exemples : Mytilus (moule), Pecten (coquille St Jacques).
4 L’ordre des Métabranches (ou Eulamellibranches).
On trouve des filaments directs et réfléchis mais soudés. Les liaisons entre les filamentsconstituées de ponts conjonctifs : on a une rigidification de la structure. sont plus importantes et
Le manteau présente une ou deux soudures ventrales (un orifice inhalant et un orifice exhalant). Exemples : Coque, Palourde.
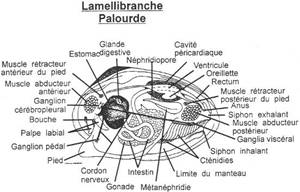
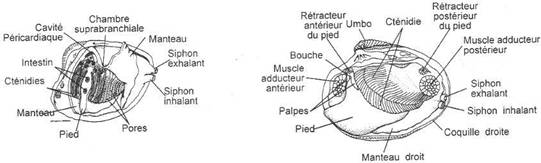
5 L’ordre des Septibranches.
Les branchies sont réduites et forment une sorte de membrane percée de pores où passe le manteau et la masse viscérale. On détermine une « chambre branchiale ».
G La classe des Céphalopodes.
Les céphalopodes sont les mollusques les plus différenciés. Ils sont souvent considérés comme les invertébrés les plus évolués. Leur système nerveux est concentré et les yeux sont de type camérulaire (semblables à ceux des vertébrés). Ce sont toutes des espèces marines. Leur taille est variable (de quelques centimètres à plusieurs mètres). Les plus grands sont les calmars géants et ils sont en concurrence alimentaire avec les gros vertébrés comme le cachalot…
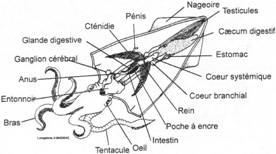
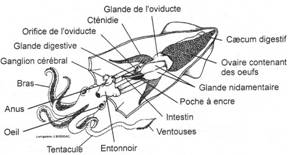
1 Organisation des céphalopodes.
Cf. TP. Exemple de la Sèche.
2 La sous-classe des Tétrabranchiaux.
Les tétrabranchiaux furent un groupe important au Primaire. Maintenant, ils ne sont plus représentés que par un genre : Nautilus.
Sa coquille est enroulée sur un plan. Elle est divisée en loges, séparées par des cloisons. L’animal vit dans la dernière loge. Il existe un siphon qui traverse toute les loges pour se terminer dans la loge initiale. Ce siphon contient un ligament qui va rejoindre la dernière loge.
Ils ont quatre branchies, quatre oreillettes, quatre reins. Ils présentent entre 40 et 50 tentacules rétractiles.
On les trouve dans l’océan Pacifique et dans l’océan Indien.
3 La sous-classe des Ammonoïdés.
Cette sous-classe est uniquement présente au Secondaire, après, ils disparaissent. Leur coquille est traversée par un siphon. Ils avaient deux ou quatre branchies.
4 La sous-classe des Dibranchiaux.
Les dibranchiaux possèdent deux branchies, deux oreillettes, deux reins.
a L’ordre des Décapodes.
Les décapodes ont dix bras péribuccaux et une coquille interne. Le manteau forme deux replis latéraux qui donnent une nageoire de chaque côté de l’animal. Exemples : Sèche, Sépia, Calmar.
b L’ordre des Octopodes.
Les octopodes ont huit bras. Ils sont dépourvus de coquille. Le manteau est sous les replis latéraux. Ce sont des animaux benthiques. Exemple : la Pieuvre.
g L’ordre de Bélemnitoïdés.
Les bélemnitoïdés sont un groupe fossile qui présentait une coquille externe.
Chapitre 5-1 :
Les Arthropodes, Généralités.
I Caractères généraux.
Les animaux de la lignée protostomienne ont subit des modifications considérables avec des spécialisations selon les milieux colonisés. Les arthropodes sont les mieux céphalisés avec les céphalopodes.
Ces arthropodes présentent des caractères d’annélides comme une chaîne nerveuse ventrale et une segmentation apparente. Ils ont aussi des caractères de mollusque avec par exemple, un cœlome réduit et un appareil circulatoire ouvert.
Les arthropodes :
- Sont des protostomiens segmentés, équipés à l’origine, d’une paire d’appendice par segment. Certains de ces appendices disparaissent ; d’autres se spécialisent.
- Montrent des formes primitives avec des appendices post-oraux tous égaux. Les formes « modernes » vont présenter des appendices spécialisés en fonction de leur position sur le corps.
- Ont une cuticule externe qui recouvre tout le corps : c’est un exosquelette rigide qui oblige la croissance par mue(s).
- N’ont des vésicules cœlomiques que chez les embryons précoces : elles vont régresser chez l’adulte alors que l’hémocœle prend de l’importance. L’hémocœle est la cavité générale où circule le sang.
- Possèdent un cœur contractile, formé à partir d’un vaisseau dorsal. Ce vaisseau est placé dans un sinus péricardique et le sang pénètre dans le cœur par des orifices latéraux paires : les ostioles.
- Ont un système nerveux bâtit sur le plan annélidien (la chaîne nerveuse est ventrale, reliée au « cerveau » par le collier péri-œsophagien). Le cerveau est hautement différencié et divisé en trois vésicules : protocérébron, deutérocérébron, tritocérébron.
- Ont leurs mouvements qui ne dépendent que de la musculature animant ces appendices locomoteurs, rattachés à l’exosquelette.
- Ont des œufs très riches en vitellus et ne présentent pas de larve trocophore (ce qui est le cas des annélides et des mollusques).
Le système excréteur néphridien des annélides a disparu.
Les arthropodes présentent 7/8ème des espèces vivantes. Ces points évolutifs fondamentaux ont été déterminant dans leur succès écologique et leur réussite.
II La diversité des arthropodes.
Chez les arthropodes vrais, on trouve trois sous-embranchements : les Trilobitomorphes, les Chélicérates, les Antennates (ou Mandibulates).
Cet embranchement est représenté dès le cambrien (-560 millions d’années) avec des formes aquatiques.
A Le sous-embranchement des Trilobitomorphes.
Ces trilobitomorphes sont des arthropodes primitifs, aquatiques, avec des antennes pré-orales et des appendices biramés sur tous les métamères. Aucun trilobitomorphe n’a atteint l’ère secondaire. On compte 3950 espèces.
B Les Chélicérates.
63000 espèces sont actuellement connues. Elles n’ont pas d’antennes. La première paire d’articles est en avant de la bouche et forme des pinces préhensiles : les chélicères. On y trouve aussi une paire d’appendices tactiles : les pédipalpes.
Le corps est divisé en deux régions ou deux tagmes :
- Un prosoma, antérieur, portant les chélicères, les pédipalpes et tous les segments munis de pattes marcheuses.
- Un opistosoma, portant des appendices réduits ou bien absents.
Les chélicérates sont divisés en trois classes :
· Les mérostomes : leur respiration est branchiale, aquatique. Ce sont les limules (ou xiphosures). Le corps est recouvert d’une carapace dorsale.
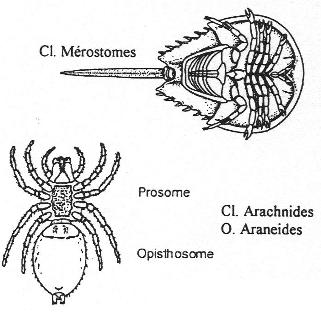
· Les arachnides : la respiration est aérienne et ils sont majoritairement terrestres. On en connaît 15 ordres et les plus connus sont les scorpions, les araignées, les opilionides ou les acariens.
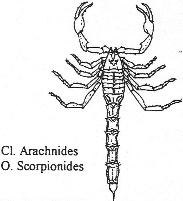
· Les pycnogonides : ce sont les « araignées de mer ». Ce sont des formes aquatiques à respiration cutanée. Le prosoma est très développé alors que l’opistosoma est très réduit.
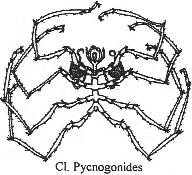
C Les antennates (ou mandibulates).
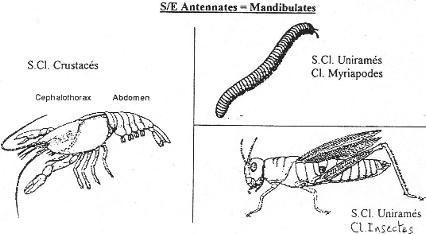
Les antennates ont des antennes et des mandibules. Ils regroupent les crustacés et les uniramés (myriapodes + insectes). La différence entre les sous-groupes est surtout faite à partir des appendices céphaliques.
1 La Super-classe des Crustacés.
Les crustacés sont aquatiques, à respiration branchiale. Ils possèdent deux paires d’antennes. On en connaît 40 000 espèces réparties en 10 classes.
2 La Super-classe des Uniramés.
Cette super-classe comprend près d’un million d’espèces. Elles sont pourvues d’une seule paire d’antennes. Les appendices locomoteurs n’ont qu’une rame.
a La classe des Myriapodes.
On trouve plus de trois paires d’appendices locomoteurs.
b La classe des Insectes (ou hexapodes).
On trouve trois paires d’appendices locomoteurs.
III Les Pararthropodes, groupe associé aux arthropodes.
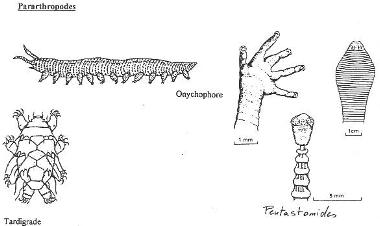
Les pararthropodes regroupent trois classes : les tardigrades, les onychophores et les pentastomides. Ces derniers seraient des arthropodes vrais déformés par le parasitisme.
Les tardigrades et les onychophores sont beaucoup plus énigmatiques. Ils possèdent des caractères d’arthropodes comme une segmentation, une cuticule, une croissance par mue. Ils présentent toutefois des particularités pouvant être considérées comme des régressions : les tardigrades n’ont ni antennes, ni pièces buccales, ni système respiratoire. Les appendices articulés peuvent être présents ou non.
Le problème de l’intégration de ces trois classes aux arthropodes n’est pas résolu.
IV Signification fonctionnelle de la métamérie.
Comme chez les annélides, le corps est à symétrie bilatérale, avec une succession de segments : les métamères. Il y a segmentation ganglionnaire du système nerveux.
A La métamérie des arthropodes.
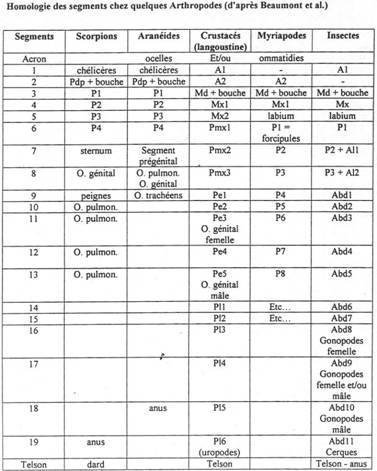
Chaque segment est entouré de plusieurs plaques (sclérites) et sont unis (les segments) entre eux et aux métamères voisins par une membrane ventriculaire.
On trouve un tergite dorsal, un sternite ventral et deux pleurites ventraux.
Un métamère est traversé par : le tube digestif, le cœur (vaisseau contractile dorsal), la chaîne nerveuse, des faisceaux musculaires longitudinaux dorsaux et ventraux.
La musculature circulaire a disparu. A la série des métamères s’ajoutent :
- L’acron dans la région antérieure, qui porte le protocérébron.
- Le telson, à l’arrière, qui porte l’anus.
Cette métamérie hétéronome résulte de la différenciation de régions (les tagmes). Chaque tagme comprend une série de métamère pour la même fonction.
Les appendices des tagmes sont spécialisés dans des différentes fonctions. Les arthropodes primitifs ont un grand nombre de segments, pouvant être indéfini, alors que les formes spécialisées ont un nombre de segments réduit et fixe. Il y a donc une division du travail entre les différents groupes de segments. La région antérieure est sensorielle et masticatrice ; la région moyenne est locomotrice ; la région postérieure porte les organes génitaux.
B Signification fonctionnelle de la métamérie.
La segmentation chez les cœlomates semble avoir une double signification : collecte de nourriture et locomotion. Chez les annélides, la segmentation est d’abord liée à la locomotion : toute réduction de la mobilité s’accompagne d’une altération de la métamérie. Par exemple, les annélides sédentaires ont une métamérie altérée par rapport aux annélides errantes.
Chez les arthropodes, on pense que la segmentation est d’abord liée à la collecte de la nourriture. Cela est basé sur un groupe de crustacés, soit fixés, soit parasites : les cirripèdes. Chez les formes fixées, on trouve deux types morphologiques :
- Les Lepas, à aspect de fleur composée, avec une carapace de plusieurs plaques calcaires (le capitulum) et un pédoncule de fixation.
- Les Balanes, dépourvus de pédoncule mais protégés par une muraille calcaire.
Dans les deux cas, le corps a six paires d’appendices troncaux en forme de fouet. Les cirres sont munis de soies plumeuses pour la capture du plancton. La sédentarité n’affecte pas la segmentation, probablement, car celle-ci est liée à la collecte de nourriture, d’où, les six paires d’appendices.
L’altération de la métamérie ne peut donc être associée qu’à un changement radical des habitudes alimentaires, comme chez les parasites. C’est le cas chez les cirripèdes rhizocéphales comme la Sacculine.
à La segmentation n’est que secondairement associée à la locomotion qui se fait dans un premier par nage appendiculaire.
V La céphalisation.
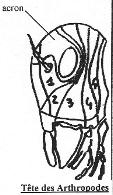
La tête résulte de la fusion de l’acron avec les cinq premiers métamères. L’acron porte les ocelles et les yeux composés. Les métamères 1 à 5 portent les appendices spécialisés dans les fonctions sensorielles (exemples : antennes et antennules) et/ou dans les fonctions masticatrices (exemples : mandibules, pattes-mâchoires).
Les métamères 1 et 2 sont en position pré-orale et leurs centres nerveux (deuto et tritocérébron) vont fusionner avec le protocérébron pour former le cerveau.
Les métamères 3 à 5 sont post-oraux et leurs ganglions fusionnent en un ganglionsous-œsophagien innervant les pièces masticatrices.
Ces deux ensembles sont liés par un collier péri-œsophagien. La tête des arthropodes reste controversée sur plusieurs points. Les formes primitives étaient proches des crustacés céphalocarides : une tête bien individualisée avec un acron plus cinq métamères. La tête est séparée du premier métamère thoracique par une limite inter segmentaire. On admet que cette disposition représente la structure fondamentale des arthropodes. Cette structure fondamentale est conservée chez les myriapodes avec une meilleure individualisation, grâce à un cou. C’est la même chose chez les insectes.
Toutefois, la tête de nombreux crustacés et de tout les chélicérates s’éloigne de ce type fondamental. Les crustacés ont une céphalisation qui fait intervenir les deux premiers segments du tronc qui vont se souder à la tête primitive. La partie dorsale de la carapace recouvre plus ou moins l’ensemble du corps. Les premiers appendices thoraciques forment des pattes-mâchoires (PMX) venant participer à la mastication. La tête est une unité anatomique mais aussi fonctionnelle. Elle regroupe les éléments pour les fonctions nerveuses (et sensorielles) et de nutrition.
Chez les chélicérates, la partie antérieure est longue, d’un seul tenant. De l’avant vers l’arrière, on trouve une paire de chélicères, une paire de pédipalpes (deuxième métamère plus bouche) puis quatre paires locomotrices. Le premier segment est dépourvu d’appendice. Le premier métamère porte les chélicères. Tout cet ensemble peut être assimilé à une tête primitive mais il assure aussi la fonction locomotrice.
On doit admettre que les pattes locomotrices correspondent à des segments du tronc ayant fusionné avec la tête ; l’ensemble formant le prosoma.
Chapitre 5-2 :
La locomotion chez les Arthropodes.
Connaître les mécanismes de la locomotion permet de comprendre les origines d’un groupe animal : tout ancêtre supposé devra fonctionner de la même façon. La locomotion nécessitesystème squelettique transmettant une force générée par l’activité musculaire. Chez les arthropodes, c’est l’exosquelette associé à la musculature qui remplie ce rôle. toujours un
I L’exosquelette et l’appendice locomoteur.
A L’exosquelette.
L’exosquelette permet d’avoir des appendices articulés : les arthropodes ont un avantage locomoteur sur tous les animaux vermiformes, d’où, leur succès écologique. Ce squelette procure la rigidité nécessaire au mouvement. Il fournit les points d’appuis pour les mouvements de leviers et fournit des points d’ancrage pour les muscles.
Les appendices permettent le déplacement de l’animal sans avoir à utiliser toute la musculature, comme chez les animaux à squelette hydrostatique type annélides.
Chez les insectes ptérygotes, la cuticule donne les ailes, ce qui permet un déplacement sur de grandes distances. Par exemple, certaines libellules sont capables de traverser la méditerranée.
La paroi du corps comprend la cuticule, l’épiderme et une membrane fondamentale. Toutes les couches musculaires circulaires, diagonales des annélides se sont transformées et se sont dispersées pour constituer la musculature spécifique qui actionne les articulations du corps.
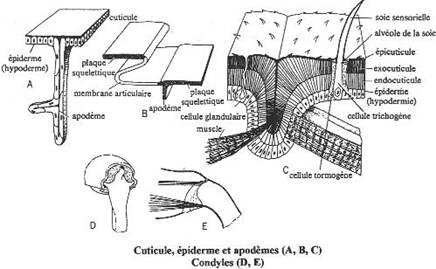
La cuticule est formée de trois couches :
- La couche externe ou épicuticule. Cette couche est très fine, imperméable, inextensible, constituée de lipoprotéines associées à des cirres et des composés phénolés limitant les pertes d’eau.
- La couche moyenne ou exocuticule. Cette couche est souvent pigmentée et forme des plaques squelettiques (les sclérites) qui sont constituées de scléroprotéines associées à de la chitine.
- La couche interne ou endocuticule. Cette couche est continue, flexible et forme les membranes articulaires entre deux sclérites successives. Elle est essentiellement composée de chitine associée à des polysaccharides.
Il existe également des structures fines aux endroits où l’épiderme s’invagine en formant des apodèmes.
Les sutures mobiles pourront montrer deux types d’adaptation. Les sclérites protecteurs se chevauchent alors que l’endocuticule forme une membrane articulaire repliée vers l’intérieur du corps. L’articulation des appendices est différente : s’il n’y a qu’un repli de l’endocuticule, tout le poids du corps reposerait sur une structure fragile : la surface d’articulation va être renforcée par des points de contact à les condyles.
Face aux condyles, la membrane articulaire (plus souple) assure la jonction entre les deux éléments squelettiques. La membrane ne sert qu’à refermer les espaces entre les deux articles.
B Les appendices locomoteurs.
Les articulations des pattes comprennent entre un et trois condyles. Leur forme et leur position vont déterminer la liberté de mouvement de l’appendice.
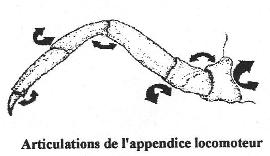
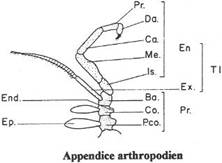
Sur l’ensemble de l’appendice, les articulations ne permettent que des mouvements de flexion et d’extension : ce sont des mouvements de type charnière. Seule l’articulation de la base est capable de rotation ; elle est responsable du mouvement vers l’avant.
Le corps des arthropodes est suspendu entre les appendices ; la marche n’implique ni élévation ni abaissement du centre de gravité. Les muscles qui animent les appendices sont situés à l’intérieur du corps.
La structure est uniforme. L’appendice est constitué d’une série d’articles qui s’affinent en allant vers l’extrémité. Les articulations fonctionnent chacune comme une charnière mais sont orientées dans des plans différents. Ce système permet à l’extrémité de l’appendice de se déplacer dans toutes les directions.
L’appendice est biramé ; il comporte une région basale : le protopodite. Cette région est constituée de trois articles : le pré-coxa, le coxa et le basis.
On retrouve ces trois articles chez les crustacés mais les insectes ne conservent que le basis qui sera appelé « la hanche ». Dans ce cas, on suppose que les deux articulations précédentes se sont intégrées au pleurite.
A partir des protopodites se sont développés des expansions, les épipodites, à rôle surtout respiratoire.
Ensuite on trouve une rame externe, l’exopodite, qui prend une fonction respiratoire en se transformant en branchies. Cette expansion n’existe que chez les crustacés et les trilobites.
Finalement, on a une rame interne, l’endopodite, qui est composée de cinq articles :
II La locomotion chez les crustacés.
A Les crustacés primitifs.
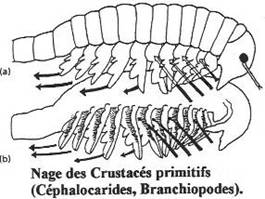
Chez les crustacés primitifs, représentés par les léphalocarides et les branchiopodes, les segments thoraciques portent des appendices qui servent à la fois à la nutrition et à la locomotion.
L’appareil locomoteur agit comme une pagaie et la surface présentée à l’eau est maximale à l’impulsion et minimale au reploiement.
La nage est favorisée par la contraction des espaces formés entre les appendices. L’eau est aspirée entre les appendices dans la région antérieure puis est expulsée latéralement dans la région postérieure. Le mouvement d’eau est unidirectionnel et ne peut s’inverser car, une fois que l’eau est aspirée dans la région antérieure, les exopodites (externes) vont jouer un rôle de valve.
Les particules sont captées au passage par les soies de l’endopodite ; elles passent ensuite vers une gouttière médio-ventrale, puis sont dirigées bers la bouche.
à Les appendices servent à la fois à la locomotion et à la nutrition : ils ne sont que peu spécialisés. Les crustacés sont uniquement nageurs.
B Les crustacés supérieurs : les malacostracés.
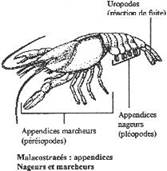
Chez ces crustacés, on observe le passage à la marche benthique, ils conservent quand même une capacité à nager. Ils conservent à la fois des appendices marcheurs thoraciques (les péréiopodes) et des appendices nageurs (les pléopodes).
Une tendance évolutive peut conduire à une réduction, voir même une perte, de la capacité à marcher : c’est le cas des Eucarides. D’autres espèces peuvent perdre la capacité à nager, par réduction ou disparition des pléopodes : cas chez les décapodes Brachyoures (crabe) et les Macroures (langouste).
Chez les macroures, les pléopodes sont réduits et ne peuvent pas servir à la nage. Les animaux ont développé une rame caudale, formée du pléopode 6 et du telson. Un retour brutal de cette rame permet la réaction de fuite vers l’arrière.
Chez les crabes, l’abdomen est très réduit. Ces animaux marchent sur le fond grâce à leurs péréiopodes. Il y a quelques exceptions car certains crabes nageurs (genre Callinecte) ont modifié la forme du péréiopode 5 qui devient aplati à son extrémité. La nage est réalisée par la rotation de ces péréiopodes.
III La locomotion en milieu terrestre.
Les arthropodes sont plus denses que l’air et leur exosquelette va interagir avec le support. Les appendices supportent des forces de flexions orthogonales à leur axe et des forces de torsion qui agissent dans l’axe de l’appendice.
D’un point de vue physique, la structure qui supporte le mieux ces contraintes est un cylindre creux.
On reconnaît trois types de locomotion : la marche, le saut, le vol.
A La marche.
La marche implique une rotation axiale de l’appendice, réalisée par l’article basal (hanche). On distingue quatre phases :
- Elévation de l’appendice.
- Mouvement vers l’avant.
- Abaissement.
- Rétraction vers l’arrière du corps (l’extrémité de la patte reste en contact avec le sol). L’animal se déplace en avant de ce point fixe.
C’est le jeu des diverses articulations de l’appendice qui maintien le corps à la bonne hauteur.
Chez les myriapodes, on distingue les Diplopodes, herbivores (mille-pattes) et les Chilopodes, carnivores (Scolopendre).
Chez les diplopodes, il y a un grand nombre d’appendices courts, chacun entrant vers l’avant juste après celui qui le précède. Il se forme une onde locomotrice dirigée vers l’avant, qui ne déplace qu’une patte à la fois. Ce mécanisme ne permet qu’une locomotion très lente car une force assez importante s’exerce vers la patte antérieure pour obtenir une vitesse faible. Le type de locomotion convient à ces animaux herbivores qui vivent souvent dans le bois en décomposition ou dans la terre meuble.
Les chilopodes (prédateurs actifs), doivent être capables d’une grande vitesse de déplacement. Plus la vitesse augmente, plus le nombre d’appendices en contact avec le sol va diminuer et les ondulations du corps s’amplifient. Le but des ondes est d’accroître l’enjambée. Ce type de locomotion demande une grande dépense énergétique vu que le corps est mis en jeu. Le problème est résolu par l’accroissement de la longueur des pattes et par la réduction du nombre de pattes. C’est une tendance évolutive très marquée chez les arthropodes.
Les crustacés décapodes ont cinq paires de péréiopodes mais 3 ou 4 servent à la marche. Les autres sont transformés en pinces.
Les arachnides ont quatre paires de pattes. Les insectes ont trois paires.
La trajectoire de chaque appendice est différente pendant la marche et ces trajectoires ne se recoupent jamais.
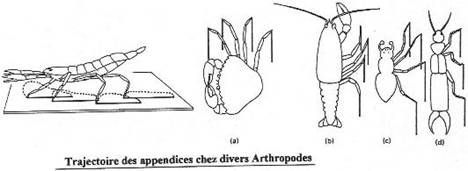
Moins il y a de pattes, plus il est difficile d’obtenir la stabilité.
Chez les hexapodes, la séquence de marche fait qu’il y a toujours trois pattes en contact avec le sol, formant ainsi un trépied. Le centre de gravité de l’insecte se projette toujours dans le triangle formé par les extrémités des pattes.
Les crabes se déplacent latéralement. La traction est réalisée par les articulations inférieures de l’appendice.
B Le saut.
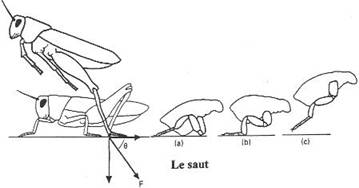
Certains insectes présentent une adaptation au saut qui est avant tout une réaction de fuite. Pour sauter, un insecte doit exercer une force sur le support afin d’obtenir une vitesse de décollage qui soit en accord avec sa masse.
½ x m x v² = m x g x h.
v est la vitesse au décollage, m est la masse de l’insecte.
H = v² / 2g ; h = Ec / m x g.
La force transmise aura une composante verticale et une composante horizontale. La résultante donne la direction du saut. L’accélération continue tant que les pattes sont en contact avec le sol. C’est pour cela que les insectes sauteurs ont des pattes III allongées ; elles augmentent le temps d’action de saut et contribuent à accroître l’accélération.
C Le vol.
La cuticule va jouer un rôle important, elle comprend les propriétés idéales pour former une aile.
C’est toujours une membrane fine, à la fois légère et flexible, supporté par des nervures chitinisées. Les insectes ailés sont des ptérygotes. On distingue deux types d’adaptation structurale.
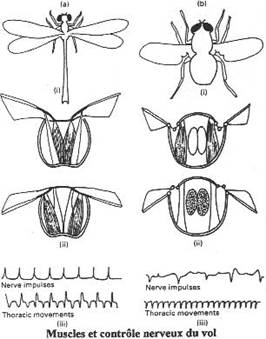
Chez les formes primitives comme les Odonates (Libellules), les ailes sont animées par des muscles directs : ils sont directement rattachés à la base des ailes. Il existe une relation entre les impulsions et les mouvements d’air : une contraction provoque un battement. On observe entre 20 et 40 battements par seconde, le contrôle nerveux étant synchrone.
Chez les formes évoluées (la majorité), le mouvement des ailes est obtenu par l’action de muscles indirects qui vont s’attacher aux sclérites thoraciques. Ces muscles changent la forme du thorax, ce qui provoque les battements. Les ailes sont articulées sur le thorax. Le moustique peut atteindre 2 218 battements par seconde. Les muscles jouent sur les propriétés élastiques du thorax. Le contrôle nerveux est asynchrone. Une contraction déclenche donc plusieurs battements.
Le mécanisme du vol.
Le vol est induit par des flux d’air, générés par les battements d’ailes. Les battements induisent des vortex (masses de fluide en rotation annulaire). Le vortex est ensuite rejeté vers l’arrière avec une direction donnée et libère de l’énergie.
Le battement des ailes génère la circulation de l’air à partir de la surface ; l’air est rejeté vers l’arrière quand l’aile se reploie ou simplement quand elle change d’inclinaison.
L’insecte est propulsé par la force de réaction qui résulte de l’expulsion du vortex.
Exemple des différentes phases de vol du décollage d’un papillon.
1- Les ailes sont accolées en position haute.
2 et 3- Les ailes s’écartent et une circulation d’air est induite sur le bord d’attaque et est dirigée vers la face dorsale.
4- La masse d’air accélère, ce qui aspire l’insecte qui décolle du support.
5 et 6- Les ailes atteignent la position basse et l’air en circulation est projeté vers le bas sous la forme d’un vortex qui va à la fois soutenir et propulser l’animal.
Les insectes capables de vol présentent un avantage écologique : ils se déplacent très rapidement sur de grandes distances. Ils pourront coloniser des aires très vastes. Certains acquièrent des capacités de migration (recherche de conditions favorables).
Chapitre 5-3 :
La respiration chez les Arthropodes.
La respiration est l’étude des mécanismes et des structures grâce auxquelles l’oxygène est capté. Les arthropodes ont colonisé tous les milieux (aquatiques et terrestres). Il faut donc considérer deux situations différentes. Les animaux vont prélever de l’oxygène soit sous forme dissoute dans l’eau, soit sous forme gazeuse dans l’air. Il existe deux problèmes fondamentaux :
- Problème pour les animaux vivant dans les zones de balancement de marées (crabes)
- Retour au milieu aquatique sur une partie du cycle de développement (larve aquatique et adulte terrestre).
I Les arthropodes aquatiques stricts.
Dans l’eau, l’oxygène est présent sous forme dissoute. Il provient d’une part de la production par des autotrophes et d’autre part, de la diffusion depuis la surface. Ce gaz est présent en faible concentration. Les structures les plus efficaces pour capter l’oxygène sont les branchies. Ce sont toujours des expansions du tégument très fines, dont le but est d’augmenter la surface d’échange avec le milieu extérieur.
Même chez les arthropodes, des branchies devront toujours répondre à trois exigences :
- La surface d’échange doit être rapport avec la consommation d’oxygène.
- La barrière entre le milieu intérieur et le milieu extérieur doit être amincie à épithélium branchial mince.
- L’animal doit mettre en œuvre une convection externe : la ventilation. Il y a création de mouvements d’eau continus autour des branchies. Il doit aussi y avoir une circulation sanguine qui permet le transport de l’oxygène.
è Ces problèmes sont résolus par les crustacés : ce sont des arthropodes à respiration branchiale.
A Les crustacés : organes et mécanismes respiratoires.
L’exosquelette forme une barrière à la diffusion des gaz, sauf chez les formes de petite taille comme le zooplancton où la cuticule est très mince et permet des échanges à son travers.
L’épiderme ne porte pas de cils. La ventilation branchiale ne peut pas être réalisée par des courants ciliaires typiques des invertébrés.
On constate une augmentation des surfaces spécialisées (branchies) qui vont progressivement devenir internes avec pour étape ultime, la cavité branchiale des décapodes. Les branchéostèges sont des expansions latérales de la carapace. La ventilation est assurée par le jeu d’appendices spécialisés ou non.
On distingue trois types de respiration.
1 La respiration palléale.
Ce type de respiration est rencontré chez les crustacés primitifs (Ostracodes) et chez les formes fixées (Cirripèdes). Le corps est enfermé dans la cavité délimitée par un repli tégumentaire latérale qui va former un manteau comparable à celui des mollusques.
L’hématose (oxygénation du sang) se fait directement au travers de la face interne du manteau. Ce dernier prend le rôle de branchie.
Chez les cirripèdes, la ventilation est assurée par les mouvements de va et vient thoraciques liés à la prise de nourriture (activité trophique).
Chez les ostracodes, la face interne de la carapace qui joue le rôle de branchie (elle est mince et vascularisée). La ventilation est assurée par l’exopodite (ou lame branchiale) de la Mx1.
2 La respiration appendiculaire.
Ce type de respiration est trouvé chez les crustacés branchiopodes dits « phyllopodes » (ex : Artenia).
Les branchies sont localisées au niveau des appareils locomoteurs, ce qui permet à la fois la ventilation et l’extraction de l’oxygène (plus locomotion et nutrition).
C’est un dispositif qui ne permet pas la marche benthique vu que les surfaces branchiales sont vulnérables. Ce sont donc des formes nageuses.
3 La respiration branchiale.
Cette respiration fait intervenir l’apparition de courants ventilatoires propres aux branchies et à la cavité branchiale.
Les branchies les plus simples sont observées chez les amphipodes. Ce sont simplement des différenciations des épipodites et elles ne bénéficient d’aucune protection. La ventilation est assurée par le mouvement des appendices eux-mêmes.
Chez les malacostracés, on trouve de véritables branchies distinctes des appendices locomoteurs. Elles y restent quand même associées par le fonctionnement. Les branchies sont protégées par un repli du tégument : le branchiostège. Ce dernier va former une cavité branchiale qui est soustraite aux courants. La ventilation sera assurée par les épipodites et les exopodites des appendices antérieurs.
Le dispositif le plus complexe est présent chez les décapodes.
B L’appareil respiratoire des décapodes.
1 Les branchies.
Les branchies sont directement associées aux péréiopodes et peuvent s’insérer à trois niveaux différents :
- sur le coxa (article basal) : on parle de podobranchies ;
- sur une membrane articulaire, entre l’appendice et le corps : ce sont les arthrobranchies ;
- sur un pleurite : ce sont des pleurobranchies.
Chaque branchie possède un axe vasculaire (le raphé) qui porte un ensemble de ramifications constituant une surface d’échanges. Sur le bord interne, on trouve des vaisseaux efférents alors que sur le bord externe, on a des vaisseaux afférents.
Le sang est collecté dans le corps par un sinus ventral puis, amené aux branchies. A la sortie de ces dernières, le sang oxygéné passe dans la cavité péricardique.
Il existe deux types morphologiques de branchies : les trichobranchies (Homard) et les phyllobranchies (Crabe).
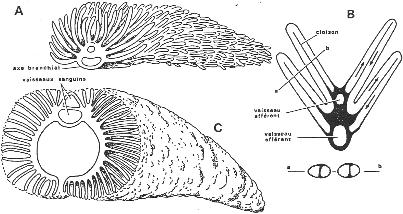
Pour les trichobranchies, les ramifications sont des filaments disposés de part et d’autre du raphé : « brosse ». Chaque filament est divisé par une cloison axiale, déterminant le trajet en « U » du flux sanguin dans les filaments.
Pour les phyllobranchies, le raphé est aplati et porte latéralement des lamelles disposées par paire symétrique. Les deux épithéliums de chaque lamelle sont écartés par des faisceaux de fibres conjonctives.
2 La cavité branchiale et la ventilation.
La disposition des branchies délimite deux compartiments : le compartiment situé entre la face interne des branchies et le pleurite est le compartiment hypobranchial, afférent. Le compartiment entre la face externe et le bronchiostège forme le compartiment épibranchial qui est efférent.
L’eau entre dans le compartiment hypobranchial en passant entre les bases des péréiopodes. Elle traverse les filaments et les lamelles des branchies et se retrouve dans le compartiment épibranchial avant d’être évacuée.
Ces mouvements de circulation sont entretenus par les battements de l’exopodite du maxille 2 (le scaphognathite). Ce dernier est également capable d’inverser brièvement son sens de battement et provoquant ainsi le nettoyage de la cavité branchiale. La fréquence des battements est fonction de la concentration en O2.
II La vie de la zone de balancement des marées.
Ces variations de marée soumettent des organismes à des exondations. Là, ces animaux ont deux solution :
- Résister à l’anoxie et se placer en anaérobiose.
- Développer des structures spécialisées leur permettant de consommer l’O2 de l’air.
è On retrouve ce dernier cas chez des décapodes comme le crabe qui présentent un appareil respiratoire permettant la consommation d’O2 dissout ou gazeux.
Cet appareil respiratoire présente trois étapes évolutives : respiration branchiale àà respiration pulmonaire. respiration tégumentaire
A La respiration branchiale.
Quand les animaux sont exondés, ils modifient l’approvisionnement en O2 des branchies. Il existe deux procédés différents :
- Aération de l’eau prisonnière dans la cavités branchiale. Ce cas existe chez les crabes de nos côtes (Carcinus maenas). C’est l’inversion du mouvement du scaphognathite qui permet l’aération de l’eau mise en réserve.
- Aération en dehors de la cavité branchiale. On trouve ça chez les crabes pompeurs de la famille des Grapsidés et des Ocypodidés. Quand ils sont exondés, ces animaux pompent l’eau par une fissure se trouvant dans la partie postérieure du corps. L’eau circule vers la cavité branchiale puis est rejetée par des orifices exhalants dans la région antérieure. Cette eau va s’écouler le long de sillons latéraux sur la carapace. Pendant ce trajet, elle se ré-oxygène et est récupérée par la fissure d’aspiration postérieure.
B La respiration tégumentaire.
Les structures de respiration tégumentaire sont chez les formes tropicales.
Chez les crabes Scopimera, les péréiopodes ont une surface élargie et déprimée. La cuticule y est très amincie et la veine drainant l’hémolymphe se capillarise et se plaque contre l’épiderme. Ce système assure 60% des besoins en O2.
Chez les pagures (genre Cœnobita), ce type de respiration tégumentaire est localisée sur la face dorsale de l’abdomen. Le principe est lui, toujours le même.
C La respiration pulmonaire.
Certains phénomènes sont rencontrés chez des crabes menant une vie particulièrement terrestre : ils présentent une réduction du nombre et du volume de branchies. Ils vont alors développer des organes respiratoires de substitution. Ces organes se localisent dans la cavité branchiale qui se transforme en un poumon.
Ce phénomène implique quatre modifications :
- Le bronchiostège se bombe dorsalement et peut ainsi délimiter une vaste chambre épibranchiale. Le compartiment pulmonaire s’isole des branchies par un repli tégumentaire et se situe dans la partie inférieure du bronchiostège.
- La face interne du bronchiostège acquière un rôle respiratoire. Elle se plisse et se hérisse de microvillosités. L’ensemble prend un aspect spongieux.
- La vascularisation des poumons implique une modification de l’appareil circulatoire : apparition de vaisseaux afférents et efférents. Dans tous les cas, la circulation pulmonaire n’est qu’une dérivation par rapport au schéma de base des arthropodes.
- La ventilation est assurée par un scaphognathite mais le rôle ventilatoire est essentiellement pris en charge par la musculature du bronchiostège : elle soulève la paroi en créant ainsi des appels d’air.
III Les arthropodes terrestres.
Ces arthropodes trouvent de l’O2 en quantité « illimitée ». Leur seul problème est de maintenir humides les surfaces d’échange (afin de faciliter les échanges gazeux). Il doit donc y avoir une lutte contre la dessiccation : appareil respiratoire d’origine tégumentaire et appareil trachéen.
A L’appareil tégumentaire.
Les appareils tégumentaires sont les plus simples, primitifs. On note une absence de ventilation. Les pertes d’eau sont mal maîtrisées. Les animaux possédant ce mode de respiration vivent en milieu humide.
Par exemple, les araignées et les scorpions possèdent au niveau de l’opisthosomapaires d’appareils respiratoires : les phyllotrachées (= poumons). Chaque poumon est une invagination du tégument dont la paroi forme de nombreux replis se disposant en lames successives. plusieurs
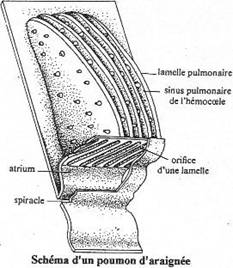
Au niveau des lamelles, la cuticule est mince et les espaces inter-lamellaires communiquent avec une cavité (l’atrium) s’ouvrant à l’extérieur par un spiracle. è Les échanges respiratoires se font par simple diffusion.
On observe des pulsations rythmiques au niveau des replis pulmonaires. Ces pulsations sont déterminées par le rythme cardiaque. C’est un phénomène typique des chélicérates
Chez certaines araignées très actives, il semble que l’approvisionnement par la voie pulmonaire soit insuffisant. Elles ont donc développé un système trachéen vrai qui amène l’O2 gazeux directement à proximité des tissus consommateurs. Ces ramifications peuvent même atteindre les pattes. Le développement de cet appareil est fonction du comportement actif de l’espèce.
B Les appareils trachéens.
C’est le système de la plupart des arthropodes terrestres et des péripattes. Cet appareil trachéen amène l’O2 gazeux aux tissus consommateurs et, approvisionnent les cellules par simple diffusion. Le sang va perdre une partie de son rôle de transporteur et ainsi perdre ses pigments.
1 La structure des trachées.
Les trachées sont des invaginations tubulaires de l’épiderme qui communiquent avec l’extérieur par un stigmate. La lumière est recouverte d’une fine couche de cuticule, en continuité avec l’exosquelette : elle est soumise aux mues.
Dans les trachées, la cuticule forme des anneaux épaissis (les taenidis) qui maintiennent ces trachées ouvertes.
Les trachées se ramifient en diminuant de diamètre. Les ramifications ultimes mesurent 3 à 5 µm et aboutissent à une cellule trachéolaire étoilée, se ramifiant en trachéole. Ce sont les extrémités aveugles des trachéoles qui sont au contact des tissus. La partie terminale de ces trachéoles est remplie d’un liquide dans lequel l’O2 se diffuse avant de pénétrer dans le tissu.
L’avantage de l’O2 gazeux est qu’il diffuse plus vite que l’O2 dissout.
2 Les appareils trachéens.
L’appareil trachéen comprend tout le réseau de trachées. On distingue deux situations :
- Chez les péripattes, araignées, diplopodes et insectes aptérygotes, les trachées sont indépendantes et peu ramifiées. Elles conservent leur disposition métamérique, chacune se terminant par un stigmate qui lui est propre.
- Chez les arthropodes les plus évolués, les trachées vont se ramifier afin d’accroître la distribution locale d’O2. Chaque trachée va se diviser en trois systèmes de distribution :
· F : Un système de ramification dorsale qui irrigue les muscles pariétaux et le cœur.
· G : Un système latéral irriguant les viscères.
· H : Un système ventral qui irrigue les muscles pariétaux et la chaîne nerveuse ventrale.
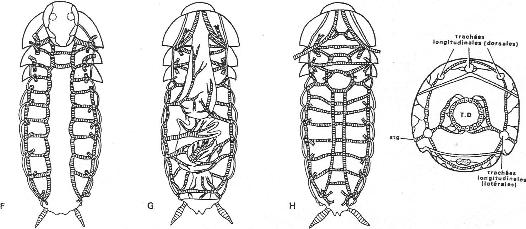
Tous ces systèmes trachéens, métamériques, sont reliés par des troncs longitudinaux et transversaux. Par conséquent, l’approvisionnement d’un organe en O2 est indépendant de la localisation des stigmates.
Les stigmates :
Chez les aptérygotes, ces stigmates sont l’ouverture directe des trachées sur le milieu extérieur. Chez les formes les plus évoluées, les stigmates s’ouvrent sur une petite cavité, l’atrium, qui assure l’isolation et la protection des conduits respiratoires. Le stigmate peut être refermé par une lèvre chitineuse mobile. Chez les fourmis, des muscles sont responsables de l’ouverture et de la fermeture des stigmates.
3 Le fonctionnement.
Le principal avantage de l’appareil trachéen est la vitesse de diffusion de l’O2 gazeux. L’élimination du CO2 se fait par les mêmes voies mais ce gaz diffuse facilement au travers des tissus : une part importante de CO2 est éliminée en passant par les trachées, puis dans l’hémolymphe puis dans le milieu extérieur.
Les mouvements ventilatoires.
Les mouvements ventilatoires améliorent la circulation en O2. Les trachées sont en relation avec des muscles locomoteurs, ce qui accroît la circulation de l’air. L’expiration est active et c’est la contraction des muscles abdominaux dorsaux-ventraux qui font diminuer le volume de la cavité viscérale et donc, expulser l’air vers l’extérieur. Les muscles longitudinaux peuvent intervenir en télescopant les métamères abdominaux.
L’inspiration est passive, résultant du relâchement de ces mêmes muscles.
L’intensité des mouvements musculaires dépend des besoins en O2. Un criquet (migrateur) pompe entre 40 et 250mL/g/h selon son activité. Une abeille a un volume maximum de ventilation de dépassant 400 fois le volume de repos.
La consommation d’O2 est à peu près constante mais le CO2 est rejeté de façon cyclique : les insectes évolués ont une respiration cyclique. La périodicité de CO2 est variable selon l’espèce (30 secondes à plusieurs heures). Entre les expirations, la quantité de CO2 rejetée est très faible. Dans le même temps, la consommation d’O2 peut être jusqu’à 100 fois plus importante.
Puisque le bilan respiratoire est déséquilibré, on peut se demander ce que devient le CO22 est consommé. Comment les stigmates permettent un flux constant d’O2 en ne laissant pas sortir d’O2 ? formé pendant que l’O
Les réponses ne sont que des hypothèses :
- L’excès de CO2 serait stocké sous forme dissoute dans l’hémolymphe et les tissus ; puis, serait brusquement libéré à l’ouverture des stigmates.
- On constante que pendant un cycle respiratoire, les stigmates sont ouverts au moment de l’expulsion de CO2, puis, se ferment hermétiquement et après quelques minutes, on observe une série de palpitations très rapides qui se prolongent jusqu’à l’ouverture suivante.
Pendant l’ouverture, la pression gazeuse trachéenne devient égale à la pression atmosphérique. Lors de la seconde phase, celle de fermeture des stigmates, la pression trachéenne diminue car l’O2 est consommé. Pendant les palpitations, la dépression du mélange gazeux provoque des appels d’air et la pression interne se rapproche progressivement de la pression atmosphérique : le flux entrant est donc faible, mais aussi constant.
La teneur en CO2 varie : environ 6,5% en fin de cycle et 3% pendant la fermeture des stigmates. La majeure partie étant dans les tissus et l’hémolymphe, sa concentration dans le mélange gazeux augmente progressivement jusqu’à l’ouverture de stigmates (décharge de CO2).
On trouve aussi un mécanisme cyclique pour l’économie de l’eau, réduisant le temps pendant lequel la vapeur d’eau est dissipée.
IV Le retour au milieu aquatique.
Même si de nombreux arthropodes se sont affranchis du milieu aquatique, un certain nombre d’espèces y reste inféodé. Ce sont surtout des insectes à adulte aérien et à larve aquatique. Par exemple, on peut citer les odonates (libellule), les éphémères, les coléoptères, les dycticidae. Dans tous ces cas, l’adulte reste proche du milieu aquatique. D’autres espèces vont être tout le temps dans l’eau.
Dans tous les cas, selon l’importance de la phase aquatique, les modifications de l’appareil respiratoire seront plus ou moins importantes : il y obligatoirement réapprovisionnement à la surface.
Certaines espèces ont des structures néoformées leur permettant la consommation d’O2 dissout : c’est alors un véritable retour à la vie aquatique.
A Utilisation de l’O2 atmosphérique.
L’appareil respiratoire n’est pas modifié ; l’approvisionnement en O2 est assuré soit par des siphons en surface, soit par des réserves emportées pendant la plongée.
1 Les siphons respiratoires.
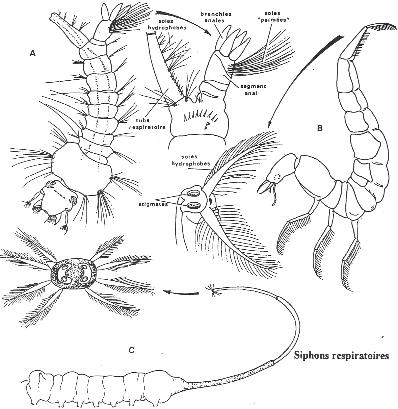
Chez les espèces à siphons, les stigmates ont disparu sauf la dernière paire abdominale. Ils sont portés par le siphon. Ces stigmates s’ouvrent à la base du siphon (Hétéroptères, Nèpes), soit à l’extrémité (larves de Culicidae [moustiques], larves de Dycticidae, larves d’Eristoles [diptères]). Chez les Eristoles, les siphons sont extensibles jusqu’à vingt fois la longueur du corps.
Parfois, les orifices respiratoires sont entourés de soies hydrophobes qui permettent la retenue de l’eau à l’extérieur.
2 Les réserves immergées.
L’animal constitue une réserves d’air sous forme d’une bulle qu’il emmène avec lui. Ce moyen de respiration est fréquent chez les adultes (Dyctidae, Gyrinidae). La bulle permet l’utilisation de l’O2 par les stigmates qui sont non modifiés. Cette bulle peut être sous les élytres ou à l’extrémité de l’abdomen.
Chez les Notonectes, la bulle est retenue par un tapis de soies microscopiques hydrophobes. Chez les Coléoptères Elmidae, les adultes sont aquatiques, vivant dans des eaux courantes, froides et oxygénées. La bulle est localisée au niveau du sternum : elle forme le « plastron » qui joue un rôle de branchie physique. Du fait de la respiration (où l’O22 diminue dans la bulle et devient inférieure à la pression en O2 dissout, donc, l’O2 dissout diffuse dans la bulle. est consommé), la pression en O
L’azote gazeux est beaucoup plus soluble que l’O2 et s’échappe de la bulle : l’animal doit alors remonter pour reconstituer sa réserve en azote.
Chez certains arthropodes, on trouve des animaux qui vivent dans des cavités remplies d’air. C’est le cas de l’Argyronète (araignée de mer) et là, la bulle est maintenue dans un tissage de soie.
B Utilisation de l’O2 dissout.
Dans ce cas, on note l’apparition de structures néoformées. Il s’agit souvent de larves d’insectes dont l’adulte est aérien. On y trouve les Ephémères, les Odonates et les Plécoptères. Le système trachéen existe chez les formes larvaires mais les stigmates sont clos.
1 Les trachéo-branchies.
Les trachées sont approvisionnées en O2 après la diffusion au travers des structures néo-formées rappelant les branchies. Ce sont de simples expansions de tégument renfermant des trachées fines et très découpées. Elles sont reliées au système trachéen comme chez les formes terrestres.
Chez les Odonates anisoptères, la surface respiratoire est localisée au niveau de la cavité rectale. La musculature rectale assure le renouvellement de l’eau par des mouvements d’aspiration et de rejet.
2 La respiration branchiale ou cutanée.
L’appareil de respiration aérienne disparaît (exemple : Hydracoriens). Dans le cas où il persiste, il sera remplie d’hémolymphe et clos. C’est le cas des Diptères chironome et simulie. L’O2 va pénétrer à travers le tégument au niveau des organes d’échanges et va diffuser dans l’hémolymphe è ce sont des « branchies sanguines ».
L’intérêt de cet appareil : les chironomes peuvent vivre dans un milieu avec une concentration en O2 très faible car ils vont synthétiser un pigment respiratoire.
Chapitre 5-4 :
Le Développement Embryonnaire des Arthropodes.
I Généralités.
Dans la majorité des cas, le jeune arthropode, au moment de l’éclosion, a un aspect larvaire. Les formes larvaires varient fortement d’un groupe à l’autre. On distingue deux types de développement embryonnaire.
A Développement de type anamorphique.
Ce type développement est aussi appelé développement dilaté. Au moment de l’éclosion, tous les segments ne sont pas formés. Les nouveaux segments s’ajouteront en avant du pygidium au fur et à mesure des mues. On peut citer l’exemple des limules (Xiphosures) dont la larve trilobitomorphe va acquérir progressivement le nombre de segments et d’appendices définitifs. Dans le cas des araignées de mer (Lycmogonides), on a trois paires d’appendices au départ ; les autres se formeront au cours des mues.
La plupart des chélicérates et des crustacés sont anamorphiques, ainsi que les insectes amétaboles (sans ailes).
B Le développement épimorphique.
Ce type de développement est direct. Le nombre de segments est définitif au moment de l’éclosion. C’est le type le plus répandu chez les insectes (ptérygotes) bien que les larves ne ressemblent pas à l’adulte.
Le développement post-embryonnaire est très riche en variations de détails. On détaillera les types les plus intéressants et les plus complexes : les crustacés et les insectes.
II Le développement post-embryonnaire des crustacés.
On passe d’un développement anamorphique (formes nageuses) à un développement épimorphique (forme benthiques). On prendra comme exemple les crustacés pénéïdes (décapodes). On distingue plusieurs stades larvaires et ce sont les mues successives qui permettent de passer les divers stades. Le schéma général est :
1-Nauplius 2-Métanauplius 3-Protozoé 4-Zoé 5-Mysis.
A Le stade Nauplius.
A l’éclosion apparaît cette larve nageuse et planctonique. Le corps mesure 0,5mm de long. C’est la forme la plus primitive chez les crustacés. Son existence n’est pas généralisée car les crustacés peuvent éclore à divers stades larvaires.
Le corps est ovoïde, sans trace de segmentation apparente. Il est prolongé par deux soies caudales et il porte trois paires d’appendices (antennes, antennules, mandibules). Le protopodite des antennes et des mandibules porte sur sa face interne, des endites à rôle masticateur.
Dans la région antérieure, on a un œil nauplien (impair) et dorsal. Celui-ci résulte de la fusion de plusieurs ocelles (deux latérales et une médiane).
Le corps, apparemment non segmenté, comprend en réalité l’acron, les segments oculaire, antennulaire, antennaire et mandibulaire et le telson.
Il existe plusieurs stades nauplius qui ne diffèrent que par l’ornementation de la cuticule et par une organisation croissante de l’anatomie. La durée de vie cette larve est de 1,5 jours. La dernière mue permet le passage au stade suivant.
B Le stade métanauplius.
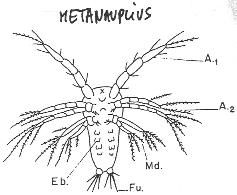
La larve a une forme de raquette. Elle mesure au maximum 0,6mm de long. Elle possède des ébauches des quatre métamères post-mandibulaires qui ont donc les bourgeons des maxillules, des maxilles et des deux premières paires de pattes mâchoires.
Le telson est pourvu d’une furca donnant deux expansions latérales.
Cette larve subit jusqu’à sept mues sans modifications majeures, le tout en 1 jour et demi.
La dernière mue permet le passage au stade suivant.
C Le stade protozoé.
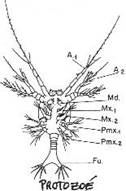
On commence à reconnaître la forme des la crevette : on a alors deux régions : une région antérieure, ovale avec carapace et une région postérieure, cylindrique, allongée, portant des traces de segmentation (furca développée).
On trouve ensuite cinq paires d’appendices céphaliques qui ont acquis leur organisation définitive. L’œil nauplien est encore fonctionnel, mais sans carapace. Les yeux composés sont pédonculés.
C’est le stade le plus long ; les mues se poursuivent sur 4 à 5 jours et la larve atteint 1,5mm.
A la fin de ce stade, les yeux sont bien différenciés et la segmentation de l’abdomen est achevée.
D Le stade zoé.
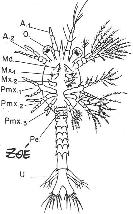
Ce stade dure trois jours et se caractérise par l’apparition de la troisième paires de PMx (pattes mâchoires). On voit apparaître l’ébauche des cinq segments du péréion.
Les segments abdominaux sont bien distincts mais seuls le dernier segment abdominal porte des appendices : les uropodes et le telson à la lame caudale.
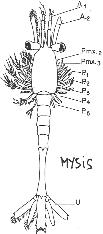

Au cours de ses mues, la larve acquière des pléiopodes. Les segments thoraciques sont différenciés et portent les péréiopodes P1 à P5. Le céphalon et le péréion se recouvrent d’une carapace non divisée : le céphalopéréion.
La dernière mue donnera l’adulte.
Les stades larvaires des groupes primitifs s’insèrent dans le développement post-embryonnaire des groupes les plus élevés.

· Chez les Mysidés (crevettes nageuses), le développement post-embryonnaire commence avec la larve nauplius et s’achève au stade zoé. L’adulte est de type mysidé.
· Chez les Péréidés, on retrouve les cinq stades, de nauplius à mysis.
· Chez les Malacostracés, comme la langoustine, il y a éclosion au stade mysis.
· Chez les Brachyoures, il y a rajout d’un stade mégalope dont la structure rappelle celle de l’adulte.
Chaque fois que les crustacés sont adaptés aux eaux douces, les stades larvaires sont abrégés et chez les écrevisses, le développement est direct.
III Développement post-embryonnaire des insectes.
A l’exception des aptérygotes, la larve possède tous ses segments à l’éclosion.
Cette larve devra subir des transformations plus ou moins importantes avant d’atteindre le stade adulte reproducteur (le stade imago).
On distingue quatre types de développement post-embryonnaire d’où quatre groupes d’insectes.
A Le développement amétabole.
On rencontre ce type développement chez les aptérygotes (Collemboles, Thysanoures). Les jeunes ressemblent aux adultes mais sont plus petits. Le nombre de segments définitifs s’acquière définitivement mais sans métamorphose. La mue d’adulte, ou imaginale, donne l’imago qui possède des gonades fonctionnelles.
Les adultes continuent à grandir par mues successives qui alternent avec des cycles de reproduction.
B Les insectes hétérométaboles paurométaboles.
Ces insectes sont des ptérygotes. On trouve différents ordres comme : les Orthoptères, les Dictyoptères, les Phasmoptères, les Dermaptères (perce-oreille) et les Hyménoptères (punaises).
L’éclosion va libérer une larve différant de l’adulte par sa taille et l’absence de pièces génitales et d’ailes. Ce cycle de développement est dit holobiontique (dans le même milieu).
Le développement est progressif : il y a augmentation de la taille et développement des pièces génitales et des ailes. Les ailes apparaissent au niveau du mésothorax et du métathorax (en position dorsale). Elles sont d’abord enfermées dans les replis du tégument (les ptérothèques). Chez les larves, les ébauches d’ailes sont visibles extérieurement : ce sont des exoptérygotes.
La mue imaginale montre une croissance accélérée des pièces génitales et des ailes on obtiendra alors un adulte qui ne mue plus et qui pourra connaître plusieurs cycles de reproduction.
C Insectes hétérométaboles hémimétaboles.
Ce type d’insecte est notamment représenté par les Ephéméroptères et les Odonates. L’éclosion va libérer une larve différant de l’adulte par sa taille, l’absence de pièces génitales et d’ailes. Toutefois, la larve se distingue de l’imago par son habitat et son mode de vie.
è Le cycle se déroule sur deux type de milieu avec souvent, une larve aquatique et un adulte terrestre : cycle amphibiontique.
La larve possède des pièces buccales broyeuses. La mue imaginale implique d’importantes modifications.
Dans tous les cas, des ébauches ailes n’apparaissent que dans le dernier stade larvaire : chez la nymphe (à insectes exoptérygotes). L’adulte ne grandit plus et peut connaître plusieurs cycles de reproduction.
D Insectes holométaboles à métamorphose vraie.
La larve est très différente de l’adulte, par son organisation morphologique et anatomique, par son habitat et mode de vie, par son régime alimentaire. Dans ce cas, on trouve les Coléoptères et les Diptères (mouches et moustiques).
Le développement des ailes est masqué et elles ne seront pas visibles en surface, avant la fin du dernier stade larvaire : ce sont des insectes endoptérygotes.
Le nombre de mues est en général fixe pour une espèce. Dans certains cas, il est soumis aux conditions environnementales (température et nourriture).
La mue imaginale donne l’adulte mais c’est une véritable métamorphose car l’animal (adulte) change complètement d’organisation, de physiologie et de milieu (par rapport à la larve). La nymphe diffère des stades larvaires par les trois mêmes critères. Avant la mue imaginale, il y aura une mue nymphale.
1 Les cinq catégories de larves.
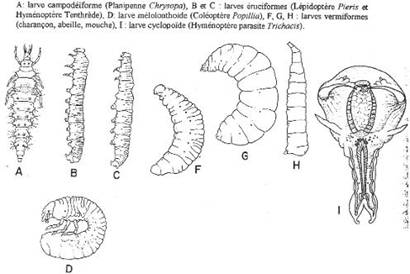
a La larve campodéiforme.
La carapace est rigide, les ocelles sont fonctionnelles et trois paires de pattes locomotrices sont présentes : c’est une larve carnassière. Cette larve existe pour de nombreux Coléoptères carnassiers (coccinelles, dytiques, carabes).
b La larve mélolonthoïde.
Ce type de larve ressemble aux larves de hanneton : elle a un corps mou, trois paires de pattes thoraciques, des pièces buccales broyeuses et pas d’ocelle. Ces larves vivent dans du bois pourri, dans des souches…
g La larve éruciforme.
C’est la larve type des chenilles de papillons. Elles ont trois paires de pattes thoraciques et n paires de pseudopodes abdominaux.
d La larve vermiforme.
C’est la larve des abeilles et des mouches. On note la disparition des appendices locomoteurs et la disparition régressive des pièces buccales et de la tête.
e La larve cyclopoïde.
On rencontre ce type de larve chez quelques Hyménoptères parasites.
2 Les nymphes.
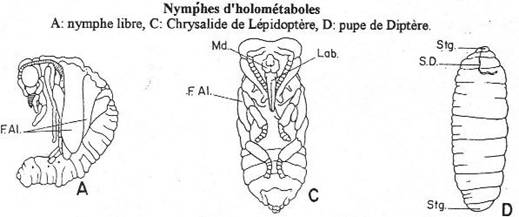
La nymphe est partiellement ou totalement immobile. L’organisation de la forme de la larve est fortement remaniée en vue de la transition vers la phase adulte.
a La nymphe libre (nue).
Les nymphes de ce type sont blanchâtres, immobiles et possèdent des appendices libres mais immobiles. On les trouve chez les coléoptères et typiquement chez Ténébrio.
b La chrysalide ou nymphes momies.
Ces nymphes sont enfermées dans un cocon de soie. C’est le cas des papillons (lépidoptères). La nymphe possède des appendices qui sont collés au corps.
g La pupe des Diptères.
Les pupes sont totalement enfermées dans la dernière exuvie de la dernière larve vermiforme. Cette larve est totalement immobile, de couleur foncée.
Dans tous les cas, la nymphe a comme l’adulte des yeux composés, des antennes, des appendices et des fourreaux alaires (replis de tégument enfermant les ailes).
La dernière mue va libérer l’adulte. Cette métamorphose débute avec la mue nymphale et se poursuit pendant tout le stade nymphal et se termine avec la mue imaginale. Durant cette période, l’insecte subit des phénomènes d’histogenèse, d’histolyse et de remaniements.
3 Histogenèse : exemple du remplacement du tube digestif larvaire par le tube digestif adulte.
Pendant le développement embryonnaire, des amas de cellules non différenciées forment des disques imaginaux et des éléments diffus (histoblastes). Au cours de la vie larvaire, les disques sont stationnaires (amas blanchâtres) et seront à l’origine des organes spécifiques à l’adulte. Ces disques s’accroissent et poursuivent leur évolution après la mue de la nymphe.
4 Histolyse.
Les organes détruits sont le tube digestif, des muscles, la trachée, des glandes thoraciques et séricigènes (pour la soie). Des hydrolases provoquent l’autolyse des cellules concernées et des cellules phagocytaires vont absorber les débris.
5 Remaniements.
Les remaniements affectent les organes communs : le cœur, certains muscles, les tubes de Malpighi (à fonction excrétrice). Les cellules subissent une dé-différenciation puis acquièrent les caractères imaginaux sans changer de destinée.
E Analyse fonctionnelle des cycles de développement.
Principe : On peut reconnaître, pendant le même cycle développement, plusieurs fonctions remplies par divers stades de développement (stades pré-adultes et adultes). On distingue différentes fonctions : une fonction de développement et de différenciation (larves) ; une fonction d’acquisition de la nourriture ; une fonction de dispersion ; une fonction d’accouplement ; une fonction de distribution de la nourriture à la progéniture ; une fonction de sélection des sites de croissance ; une fonction de ponte.
On reconnaît cinq cycles fondamentaux chez les arthropodes.
La largeur de la bande représente la quantité de ressources disponibles. Une augmentation de cet largeur montre une acquisition de cette nourriture alors qu’une diminution de cette même bande indique une utilisation de ces ressources.
En 1, on voit une transition graduelle vers l’adulte. La larve et l’adulte sont impliqués dans l’exploitation des mêmes ressources. Les adultes ont en plus les fonctions d’accouplement, de ponte et de dissémination.
En 2, on voit que les larves et les adultes se nourrissent en exploitant des ressources différentes. La larve se nourrit avec des cadavres d’animaux morts : c’est une fonction de développement et d’acquisition des ressources. On obtiendra alors une pupe qui va utiliser les ressources acquises par la larve pour son développement et sa différenciation. L’adulte va continuer à se nourrir mais à partir d’une autre source de nourriture. Ce dernier possède les fonctions d’accouplement et de sélection du site de développement des larves, de ponte.
En 3, on observe les insectes amphibiontiques (à larves aquatiques et à adultes aériens). Il y a séparation du rôle d’acquisition et de dispersion d’une manière très marquée. L’adulte ne se nourrit pas ou seulement très peu. C’est le cas des éphémères, des trichoptères et des plécoptères.
La larve est herbivore ou carnivore. Sa durée de vie varie entre quelques mois et trois ans. Elle possède la fonction de nutrition. L’adulte ne vit que quelques heures pendant lesquelles il doit s’accoupler puis pondre dans un site convenable aux larves. C’est ensuite sa mort.
En 4 et 5, on voit des adultes de certaines espèces qui remplissent les fonctions de dispersion, de ponte mais aussi de stockage et de mise à disposition des ressources sous formes provisions pour les larves.
En 4, c’est le cas des guêpes solitaires où l’adulte pond un œuf sur une proie paralysée. Cet adulte a alors remplie son rôle de dispersion, d’accouplement de sélection du cycle de ponte et de ponte. La larve doit acquérir la nourriture et la fonction de développement.
En 5, on observe les hyménoptères sociaux. Il y a un partage des fonctions assuré par une différenciation des individus en castes. Les larves utilisent les ressources mises à leur disposition pour leur croissance. Les ouvrières sont stériles et ont la fonction d’acquisition des ressources. La reine est un adulte fertile regroupant les fonctions d’accouplement, de ponte et de dispersion. Les males sont des adultes fertiles avec la fonction d’accouplement mais n’ont aucun rôle dans l’acquisition.
Chapitre 5-5 :
Prédation et Défense chez les Arthropodes.
I La Prédation.
Un prédateur est un animal qui attrape, tue et consomme d’autres animaux.
On verra la capture de proies mobiles (par des systèmes de capture) et des modalités de consommations spécialisées.
A La capture.
Les prédateurs peuvent rechercher leurs proies (scorpions), les guetter en embuscade (araignées, mantes). Tous possèdent des systèmes (armes) de capture et d’immobilisation. De plus, des soies sensorielles portées par les pédipalpes transmettent des informations afin de faciliter la capture par les pédipalpes (trichobothrie) ou les chélicères.
Par exemple, les chilopodes présentent, près de la bouche, une paire d’appendices locomoteurs qui est transformé en forcipules (à venin). La première paire de pattes est souvent devenue ravisseuse (mante).
Les systèmes de capture proviennent d’appareils pré-existant.
L’exemple des araignées.
Les araignées possèdent des glandes filières et des glandes séricigènes qui produisent de la soie (afin de former des pièges). La toile est souvent un piège passif mais peut être actif. Des araignées vont synthétiser une toile conique, tendue entre leurs pattes postérieures, qu’elles vont lâcher sur les proies.
Dans certains cas, les soies forment des sphères visqueuses qui sont pendues à l’extrémité d’un fil. Les proies seront saisies par des chélicères transformés en crochet.
Les orthognathes ont un déplacement des chélicères perpendiculaire à l’axe du corps : la taille des proies est donc illimitée. Les labidognathes ont leurs chélicères qui se déplacent dans le plan du corps, ce qui limite l’ouverture des chélicères.
B Consommation.
La majorité des insectes dont tous les arachnides, formes prédatrices, consomment leurs proies par succion. Les fluides corporels produits sont aspirés après la percée du tégument. Les arachnides ont des enzymes salivaires qui sont injectées à la proie et qui commencent la digestion de l’animal : les tissus seront liquéfiés. Pendant ce temps, la proie est tenue par les chélicères. Les sucs digestifs seront enfin ré-ingurgités.
L’appareil consommateur sera simplifié chez les acariens parasites comme les tiques (les proies sont seulement ponctionnées).
Chez les pseudo-scorpions, ce phénomène de digestion extérieure est poussé au maximum car la bouche est entourée de soies, fonctionnant comme un filtre : la proie est maintenue par les chélicères et sera entièrement dissoute. Seul l’exosquelette restera intact. Ces arthropodes réaliseront seulement une aspiration de bouillie alimentaire. Les déchets grossiers seront rejetés par les soies.
Les chilopodes et les insectes broyeurs vont triturer les aliments avec leurs pièces buccales chitineuses. Cette nourriture sera ensuite triée dans un premier intestin (antérieur) puis, digérée dans l’intestin moyen).
II La réponse des proies : défense contre les prédateurs.
Les arthropodes sont eux-même les proies de vertébrés ou d’invertébrés. Ils ont mis au point trois réponses.
A L’évitement.
La proie doit rester hors de portée du prédateur ou être invisible : migration, fuite, camouflage.
1 La migration.
La migration est caractéristique des crustacés zooplanctoniques (copépodes). Ils vont effectuer des migrations verticales où ils s’enfoncent dans une colonne d’eau et ne remontent que la nuit : ils sont peu visibles à la lumière, surtout pour les poissons qui pratiquent une chasse visuelle.
Des migrations sont observées chez des insectes aquatiques où les larves dérivent pour aller coloniser les mers en aval. Un pic de migration existe dans ce cas en début de nuit.
2 La fuite.
La fuite est une forme extrême d’évitement. Souvent, elle implique des réponses locomotrices spécialisées. Il peut y avoir utilisation de la rame caudale comme chez les dodécapodes ou utilisation des pattes 3 chez les sauteurs.
3 Le camouflage.
Chez les arthropodes, se camoufler correspond à se confondre avec son milieu.
De nombreux insectes comme les papillons ont des ailes ou un corps dont la coloration les rend invisibles sur les arbres : c’est le mimétisme de couleur.
Le mimétisme de forme se trouve par exemple chez les Phasmoptères, les Dictyoptères. Ces animaux prennent la forme de brindilles ou de feuilles.
B La dissuasion.
Cette dissuasion va être réalisée par des moyens physiques ou chimiques.
1 Dissuasion physique.
Dans ce cas, il peut y avoir utilisation de l’exosquelette qui constitue une fortification, renforcée par des expansions (exemple des crustacés qui forment des extensions de précipitations calcaires). La muraille peut être fermée par un opercule.
Dans d’autres cas, une protection externe peut être utilisée. Par exemple, les pagures protègent leur corps dans une coquille de gastéropode. En fait, les pagures réalisent une économie d’énergie en ne synthétisant pas eux-mêmes une coquille.
Il peut aussi y avoir utilisation d’un bouclier. Par exemple, le genre Cassida (coléoptère) possède un bouclier formé de matières fécales compressées. Ce bouclier est porté dorsalement par une extension de la carapace et il est orientable.
2 Dissuasion chimique.
La dissuasion chimique indirecte existe chez les pagures qui s’abritent dans des coquilles de gastéropodes recouvertes de cnidaires.
La dissuasion chimique directe est présente, par exemple, chez les insectes. Les toxines sont très souvent extraites des plantes consommées. Les Poikilocerus extraient des toxines de graminées et les stockent dans des glandes à poison avant de les projeter lorsqu’ils sont attaqués. Cette présence de toxine est souvent associée à des couleurs vives (aposématiques). Ces mêmes couleurs pourront être imitées par des espèces non-toxiques (mimétisme batésien). Toutefois, pour que cette imitation, ce mimétisme, fonctionne correctement, il faut que l’espèce non toxique soit en nombre inférieur à celui des toxiques.
C La répulsion.
Les arthropodes prédateurs peuvent devenir des proies : les organes de capture peuvent devenir des organes de défense active.
Chez les hyménoptères, l’organe de ponte (ovipositeur) est transformé en dard et ne sert plus à la ponte. Il est en relation avec la glande à venin, dans l’abdomen.
Le cas du Coléoptère bombardier (Brachinus).
Les coléoptères de ce genre projettent une sécrétion depuis une paire de glandes post-abdominales (principe actif : benzoquinone synthétisée de façon explosive). Ces sécrétions sont projetées à 100°C et accompagnées d’une détonation audible.
Le cas des Diplopodes.
Les diplopodes sont lents et donc, des proies faciles. Ils portent une paire de glandes répulsives par segment qui va sécréter du cyanure. Le fluide est libéré sous forme de jet sous pression. A chaque sac glandulaire est associé un muscle qui assure la décharge jusqu’à trente centimètres.
D L’imitation d’un prédateur.
De nombreux Lépidoptères vont pratiquer ce type de dissuasion : ils portent des ocelles sur leurs ailes qui imitent des yeux d’oiseaux.
La prédation a des modalités diverses et des réponses qui sont autant diversifiées : les phénomènes adaptatifs et évolutifs sont très poussés. Les réponses à la prédation sont très coûteuses en énergie : la croissance des populations soumises à la prédation est souvent réduite.
Chapitre 6 : L’embranchement des Cordés.
I Généralités.
L’embranchement des cordés regroupe un grand nombre d’organismes issus d’une radiation adaptative importante : un même ancêtre commun.
On distingue trois sous-embranchements :
·Les Urocordés.
Le sous-embranchement des urocordés est formé d’un petit groupe d’animaux à cycle vital dimorphique (stades adulte et larvaire sont complètement différents). Le stade adulte est fixé et le stade larvaire est libre (aquatique). Le stade larvaire est dominant dans le cycle vital.
· Les Céphalocordés ou acrâniens. Exemple : Amphioxus.
· Les Vertébrés. Des agnathes aux mammifères.
A Les caractères généraux.
Les cordés ont une symétrie bilatérale, sont métamérisés.
Ils ont tous une corde (ou chorde ou notocorde) qui est une tige endo-squelettique formée de cellules remplies de fluide, l’ensemble étant entouré d’un tissu conjonctif. Cette corde a pour rôle d’assurer une rigidité de l’animal (c’est un support longitudinal). Elle est réduite chez les vertébrés car elle est remplacée par la colonne vertébrale.
Présence d’un tube nerveux dorsal ; moelle épinière chez les vertébrés.
Présence de fentes pharyngiennes (pour la filtration chez les cordés primitifs). Ces fentes se développent de différentes façons chez les vertébrés.
Présence d’une queue bien développée en position post-anale.
B Origine de l’embranchement des cordés.
L’embranchement des cordés remonterait à 500 millions d’années (cambrien) mais les plus vieux fossiles connus sont ceux de vertébrés. On pense que l’absence de structures calcaires expliquerait l’absence de traces.
C Les vertébrés.
Les vertébrés sont caractérisés par une colonne vertébrale constituée de vertèbres métamériques. Le crâne sert à la protection du cerveau. La colonne est un support et une protection de la moelle épinière.
Classiquement, on divise les vertébrés en deux groupes : les agnathes et les gnathostomes. Ces derniers sont divisés en six groupes : les chondrichtyens, les ostéichtyens, les amphibiens, les reptiles, les oiseaux et les mammifères.
Tous les vertébrés, pendant leur développement, on une série d’organes communs :
- Le pharynx : Chez les poissons, c’est de ce pharynx que dérivent les arcs branchiaux qui donnent les branchies.
- Présence d’un ou deux reins chargés de filtrer les fluides internes. Ils servent aussi à l’excrétion.
- Présence d’un foie qui sert à la transformation, au stockage et à la détoxication des substances nutritives apportées par le sang.
- Présence d’un arc neural qui forme la partie inférieure et supérieure des vertèbres.
- Présence de deux yeux latéraux.
- Présence d’un système acoustico-vestibulaire constitué de canaux semi-circulaires, de statocystes et de la ligne latérale pour certains vertébrés.
- Présence d’un cerveau complexe comprenant le bulbe olfactif, le bulbe visuel, le bulbe auditif, les hémisphères cérébraux et le cervelet.
- Présence d’un système endocrinien développé et de différentes glandes (thyroïde, surrénales, gonades).
II Les Agnathes.
Les agnathes sont les vertébrés les plus anciens. Leur organisation est rudimentaire. Ils ont longtemps été considérés comme des « poissons ». Il en existe deux types fondamentauxLamproie et la Myxine. dont les descendants sont la
Les agnathes sont aquatiques. Leur corde est persistante. L’appareil respiratoire branchial est plus ou moins isolé du pharynx. Les branchies sont localisées dans les poches branchiales ; chacune correspondant à un métamère. Les poches sont séparées par les arcs squelettiques branchiaux.
Le squelette est rudimentaire, cartilagineux. Les agnathes présentent souvent un squelette dermique ossifié. Ils sont caractérisés par l’absence de mâchoire, de vraies dents, de ceintures, de nageoires ou de membres pairs.
A Les agnathes fossiles ou classe des Ostrachodermes.
Les Ostrachodermes possèdent une carapace sur le tégument. Leur nutrition est microphagique ; la bouche restant constamment ouverte. Ils sont regroupés en deux ensembles.
1 La sous-classe des Ptéraspidomorphes.
Ce sont les plus anciens vertébrés connus. Les plus abondants des ptéraspidomorphes étaient les hétérostracés (du silurien au dévonien). On comptait 300 espèces.
Les hétérostracés avaient un squelette externe en deux parties :
- Un bouclier céphalo-thoracique avec éventuellement un rostre.
- Une multitude de plaques recouvrant le corps un peu comme les écailles de poisson.
- On pouvait aussi trouver des épines sur ce squelette externe, quelle que soit la partie du corps.
La chambre branchiale était recouverte par le bouclier. Il y avait une ouverture sur le coté.
La queue était hétérocerque où le lobe ventral, développé, recevait le squelette axial.
2 La sous-classe des céphalospidomorphes.
Le super-ordre des Ostéostracés.
Le groupe des ostéostracés est le mieux connu des agnathes fossiles. Ils avaient une tête large et aplatie. Les yeux étaient sur la face dorsale, rapprochés l’un de l’autre.
Le bouclier était bien développé et pouvait avoir une longue corne postérieure.
La queue était hétérocerque avec le lobe supérieur plus grand que le lobe inférieur.
Les nageoires (une ou plusieurs) dorsales avaient des appendices qui faisaient penser aux nageoires pectorales.
B La classe des cyclostomes.
On a deux lignées diverses qui persistent : les lamproies et les myxines. Les deux lignées perdent le bouclier céphalo-thoracique. L’endo-squelette est cartilagineux ; le régime alimentaire microphage, au moins à l’état larvaire.
1 La sous-classe des Pétromyzonoïdes (des lamproies).
Les lamproies forment le groupe le plus nombreux des cyclostomes avec neuf genres et trente cinq espèces.
· Anatomie.
Le corps est long, cylindrique, recouvert d’une peau nue et visqueuse, riche en cellules à mucus. Les nageoires sont impaires, dans le plan sagittal du corps avec deux nageoires dorsales séparées par un court intervalle et une nageoire caudale. Ce sont de simples replis du tégument. Qui n’ont pas de rôle important dans la propulsion. Celle-ci est assurée par des mouvements ondulatoires du corps (comme chez les anguilliformes). La bouche est arrondie, antérieure et sert à la succion. Elle ressemble à un entonnoir tapissé de denticules cornés, renouvelables périodiquement.
· Le squelette.
Le squelette est composé d’une série d’arcs cartilagineux, ouverts dorsalement. C’est sur la face dorsale des arcs que passe le système nerveux.
· La musculature.
Il existe des masses musculaires disposées symétriquement le long de la corde. Elles ne sont pas divisées comme on pourrait l’observer chez les poissons où l’on a des muscles latéro-dorsaux et latéro-ventraux.
· L’appareil digestif.
L’appareil digestif forme un tube allongé et droit. Au niveau de la bouche, on trouve une langue râpeuse. La bouche est suivie par un œsophage prolongé par l’intestin qui comprend une valvule spirale.
·L’appareil circulatoire.
L’appareil circulatoire rappelle celui des poissons. Le cœur a trois cavités successives : le sinus veineux, l’oreillette et le ventricule. L’hémoglobine du sang est légèrement différente de celle des vertébrés. Elle s’apparente plus à celle des invertébrés (érythrocruorine).
·Les organes sensoriels.
On trouve des yeux primitifs, une oreille interne comprenant deux canaux semi-circulaires. La région antérieure présente dix paires de nerfs crâniens qui se détachent du cerveau.
·Biologie.
La bouche fonctionne comme une ventouse : elle s’attache à un substrat dur (pour combattre le courant) ou sert à la nutrition en se fixant sur un poisson. Les adultes sont souvent des ectoparasites de poissons et sucent leurs proies. Ce sont des carnassiers sans mâchoire. On trouve dans la bouche, des glandes spécifiques qui sécrètent des substances anticoagulantes pour avaler plus facilement le sang des proies.
Les lamproies ne se reproduisent qu’une fois dans leur vie. Le cycle est à moitié en mer, à moitié en eau douce. Les œufs donnent des larves (larve ammocète) qui vivent plusieurs années dans les sédiments des rivières. Elles subissent la métamorphose et retournent vers la mer (en automne et hiver). Elles subissent ensuite une phase de grossissement et de maturation sexuelle. Les adultes font la montaison (au début de l’hiver) et la reproduction a lieu au cours du printemps. Le cycle est appelé « migration anadrome » (les adultes, marins, se reproduisent en eau douce) (les animaux catadromes ont les adultes qui vivent en eau douce et la reproduction a lieu en mer Exemple : les anguilles).
La remontée des adultes en eau douce se fait par différents moyens : ils peuvent se déplacer seuls en nageant ou rampant ou prendre une « taxi ».Dans ce dernier cas, il s’accroche sur un poisson migrateur. Pendant la montaison, ils vivent sur leurs réserves (ne mangent pas) et s’amaigrissent. Ils se reproduisent dans des petites rivières à fonds sableux ou pierreux. Pendant l’accouplement, la femelle est fixée sur un substrat et le mâle, fixé sur la tête de la femelle. La fécondation est externe et les œufs sont fécondés au fur et à mesure de la ponte. Les adultes meurent ensuite.
Certaines espèces vivent constamment en rivière (comme la « lamproie de Planer »). Là, la phase larvaire est de longue durée (7 à 8 ans dans les sédiments). Les adultes sont juste là pour la reproduction.
La lamproie marine est consommée (Lamproie à la Bordelaise). La lamproie de Planer peut être utilisée comme appât.
2 La sous-classe des Myxinoïdes (des myxines).
Les myxines n’ont pas de parenté directe avec les lamproies. On trouve chez les myxinoïdes six genres et une quinzaine d’espèces.
· Anatomie.
Le corps a une forme proche de celle de la lamproie. On ne trouve qu’une nageoire caudale, très réduite. La peau est visqueuse, riche en cellules à mucus.
La bouche est située ventralement et est entourée de huit tentacules courts. Elle est en forme de fente.
· Les organes sensoriels.
Les organes sensoriels sont représentés par des yeux primitifs, situés sous la peau et par une oreille interne à un canal semi-circulaire.
·Le squelette.
Le squelette est beaucoup plus rudimentaire que celui des lamproies. Il ne contient pas d’éléments vertébraux.
·Biologie.
Quelques espèces sont hermaphrodites mais la majorité ont les sexes séparés. Toutes les myxines sont marines, dans les zones tempérées et froides (atlantique nord). Elles sont benthiques, vivant en grande partie enfouies dans le sédiment. Ce sont des carnivores qui vont faire des trous dans le corps de la proie pour en dévorer l’intérieur. Elles vont essentiellement s’attaquer aux poissons blessés, mourant ou morts.
C Ressemblance gnathostomes/agnathes.
Les lamproies sont beaucoup plus proches des gnathostomes que des myxines. On trouve une cinquantaine de caractères évolués chez les lamproies. Le partage de caractères évolués est appelé « synapomorphie ». Par exemple, on peut citer : présence d’une hypophyse subdivisée, innervation cardiaque, osmorégulation, présence de muscles associés aux nageoires impaires.
Les myxines ont des caractères très primitifs, comme : un système immunitaire très rudimentaire, un cœur sans innervation. Elles sont incapables de survivre à une petite modification de salinité.
L’étude synapomorphique fait penser que les lamproies et les gnathostomes ont un ancêtre commun différent de celui des myxines. Dans la classification récente (20 ans !!!) il a été créé un groupe pour les myxines et un groupe pour lamproies et gnathostomes.
III Les gnathostomes.
Les gnathostomes apparaissent à l’ordovicien. Ils partagent plusieurs caractères évolués qui attesteraient d’une ascendance commune : la myéline dans le système nerveux ; dans l’oreille interne, la présence de trois canaux semi-circulaires formant l’organe de l’équilibration ; des mâchoires qui dérivent des arcs squelettiques viscéraux 2 et 3 qui s’associent et s’articulent pour former la mâchoire et la suspension de la mâchoire sur le crâne. Les mâchoires permettent la capture de proies plus importantes. Il y a le développement de nageoires paires (qui donneront les membres) qui sont les éléments essentiels de la mobilité. La mobilité est importante pour la recherche de nourriture et de partenaire sexuel. Ces innovations font que les gnathostomes supplantent les agnathes et envahissent les eaux douces et les continents.
A Les placodermes.
Les placodermes sont les premiers gnathostomes à être apparus (silurien carbonifère inférieur). Ils avaient une plaque dermique osseuse, une mâchoire inférieure bien développée et une queue généralement hétérocerque. Ils se rapprochent du groupe actuel des chondrichtyens par la présence d’organes d’accouplement (les claspers). Ils ont vite disparu et ce sont les chondrichtyens qui les ont supplantés.
B Les chondrichtyens.
Les chondrichtyens sont les poissons cartilagineux. Les premiers représentants sont apparus à la fin du dévonien. Ils persistent de nos jours par deux groupes.
- Les sélaciens (ou élasmobranches) : ce sont les requins et les raies. Les requins ont leurs fentes branchiales latérales (pleurotrèmes) et les raies les ont ventrales (hypotrèmes).
- Les holocéphales (ou chimères).
Dans ce groupe (chondrichtyens), les principales innovations par rapport aux placodermes sont la présence d’une dentition avec des dents non soudées à la mâchoire qui sont remplacées successivement et la disparition du squelette dermique (bouclier) qui est remplacé par un squelette cartilagineux.
1 Les sélaciens.
L’appareil mandibulaire est hypertrophié. 5 à 7 fentes branchiales sont ouvertes directement à l’extérieur. Ils possèdent un spiracle (ou évent) situé en arrière des yeux. Ce spiracle permet l’entrée d’eau en direction des branchies quand la bouche est fermée ou quand les animaux mangent. Les requins sont plutôt pélagiques alors que les raies sont plutôt benthiques.
2 Les holocéphales.
Les holocéphales ont très peu évolués depuis leurs origines. Ils sont caractérisés par une mâchoire supérieure soudée au neurocrâne et des fentes branchiales recouvertes par un repli tégumentaire. Il peut y avoir fusion des dents pour former des plaques dentaires.
Ces deux groupes sont bien adaptés au milieu aquatique. Ils n’ont pas de paupières mais l’œil est en permanence humidifié et nettoyé. Les deux groupes n’ont ni oreille moyenne ni oreille externe : ils n’ont pas d’audition. Ils peuvent analyser finement les vibrations du milieu aquatique grâce à la présence de récepteurs situés sur la « ligne latérale ». Ce sont les neuromastes, situés dans des petits canaux, qui forment la ligne latérale. Les neuromastes sont composés de cellules ciliées entourées de mucus.
Les requins sont les plus spécialisés dans la réception d’ondes sonores et électriques. En effet, en plus de la ligne latérale, ils possèdent au niveau du museau, une série d’organes électro-récepteurs : les ampoules de Lorenzini. Ces organes sont ultra-sensibles à des variations de tension électrique. Ils détectent jusqu’à 10-6V de variation : ils détectent donc des tensions inférieures à celles qui existent dans le système nerveux de tout animal.
Les chondrichtyens ont une flottabilité négative (coulent quand ils sont arrêtés) à cause de la densité de leur corps qui est supérieure à celle de l’eau de mer. Il va y avoir mise en place d’un processus leur permettant de dépenser moins d’énergie pour rester à la même profondeur : accumulation de corps gras dans le foie.
Les sélaciens ont un foie qui représente 1/5ème du poids du corps alors que les poissons osseux ont un foie qui ne représente que 1/20ème du poids du corps.
L’absence de pièces osseuses permet au squelette cartilagineux d’être trois fois moins lourd.
C Les Téléostomes (acanthodiens, actinoptérygiens, sarcoptérygiens et tétrapodes).
Tous les téléostomes ont un squelette interne ossifié. Ceux qui portent des nageoires ont des rayons dermiques qui les soutiennent. Ces rayons sont constitués de lépidotriches (écailles modifiées).
1 Les acanthodiens.
Les acanthodiens n’ont pas de représentants actuels. Ce sont les premiers téléostomes et sont rencontrés dès le silurien inférieur. Ils se sont éteints entre le carbonifère et le permien.
Ils avaient des nageoires anale, dorsales, pectorales et pelvienne soutenues par une grande épine osseuse. Leur queue était hétérocerque.
2 Les actinoptérygiens.
Dans ce groupe, on trouve tous les poissons osseux sauf évidemment les acanthodiens. Ils sont apparus au silurien supérieur et sont caractérisés par la présence d’un opercule osseux qui recouvre la région branchiale. Les dents ont fusionné aux os des mâchoires. Le bord supérieur de la bouche est formé par deux os (pré maxillaire et maxillaire).
3 Les sarcoptérygiens.
On pense que les sarcoptérygiens sont les ancêtres possibles des tétrapodes actuels. Ils ont encore une forme de poisson et leurs membres ressemblent plus à des ébauches de membres de tétrapodes qu’à des nageoires de poisson. L’articulation est de type rotule, située entre la ceinture et le membre.
Ils sont bien représentés à l’état fossile mais il existe quelques lignées ayant persisté (dipneustes, crossoptérygiens).
a Les dipneustes.
Les dipneustes présentent deux types de respiration, branchiale et pulmonée. Ils ont des caractères de poisson et de tétrapodes.
Les caractères de poisson sont :
- Un système sensoriel latéral (ligne latérale)
- Un corps recouvert d’écailles
- Un système circulatoire branchial.
Les caractères de tétrapodes sont :
- Un cœur partiellement cloisonné (au niveau des oreillettes)
- Ils ont une veine et une artère pulmonaire
- La mâchoire inférieure a une suspension autostylique
- Ils possèdent un véritable coude mobile ainsi qu’un radius et un cubitus de longueur identique
- Il y a présence de deux poumons fonctionnels.
Les poumons dérivent de la vessie gazeuse. Le canal reliant les poumons au tube digestif débouche ventralement au niveau de l’œsophage. On trouve une fente : la glotte.
b Les crossoptérygiens.
b1 Les actinistiens.
Les actinistiens sont apparus au dévonien. Ils étaient bien diversifiés. Ils ont régressé et il n’existe plus qu’une forme, le cœlacanthe. Il a été découvert à la fin des années 30, au large de Madagascar. Les cœlacanthes sont caractérisés par l’absence de narines internes bien que l’on trouve deux orifices faisant penser à des choanes. Ces orifices sont recouverts par une mince couche cellulaire, qui les rend non fonctionnels.
Ils ont aussi des poumons, non fonctionnels, car ils sont calcifiés et ossifiés. Les branchies sont bien développés et la circulation sanguine est typique des poissons.
b2 Les rhipidistiens.
Les rhipidistiens n’existent que sous la forme de fossiles. Ils présentaient un crâne dont l’architecture est semblable à celle des tétrapodes. Il y a présence des maxillaires et des prémaxillaires. Le premier segment des nageoires pectorales ressemble à l’humérus des tétrapodes.
On pense qu’ils seraient les ancêtres les plus probables des tétrapodes actuels.
4 Les tétrapodes ou la sortie des eaux.
La sortie des eaux a nécessité des modifications anatomiques et physiologiques. Il y respiration pulmonée sauf chez certains animaux qui conservent des branchies ayant surtout un rôle respiratoire à l’état larvaire.
Les tétrapodes vont devoir s’adapter à la dessiccation et développer un système de régulation du milieu interne (osmorégulation) ; lutter contre la gravité (développement de membres puissants et rigidification de la colonne vertébrale). Les membres puissants servant à la locomotion voient leurs articulations se modifier (poignet pour les membres antérieurs, chevilles et genoux pour les membres postérieurs).
On va observer une adaptation au niveau de la reproduction : il y a deux choix comme stratégie de reproduction :
- La stratégie « r » : production d’un grand nombre d’œufs en espérant que quelques uns se développent. Les œufs sont sans protection.
- La stratégie « K » : cette stratégie consiste à produire un nombre d’œufs limité tout en assurant la protection de ces œufs pendant leur développement.
Les tétrapodes sont divisés en deux groupes :
- Les amphibiens
- Les amniotes.
IV Les amphibiens.
Les lépospondyles sont les amphibiens fossiles ; les lissamphibiens sont les amphibiens actuels. Ces derniers regroupent les Anoures, les Urodèles et les Apodes (ou Gymnophiones).
Les lissamphibiens ont une peau lisse. Leurs caractères généraux sont :
- Tétrapodes,
- Anamniotes,
- Peau nue et humide,
- Pentadactyles, (sauf les apodes)
- La respiration larvaire est branchiale et la respiration adulte est pulmo-cutanée,
- Le cœur est constitué de trois cavités (deux oreillettes et un ventricule),
- Le crâne s’articule sur la colonne vertébrale par deux condyles avec l’atlas.
- Le développement post-embryonnaire comporte généralement une métamorphose. Ils passent une partie de leur vie en milieu aquatique.
A Caractères de familles.
1 Les anoures (super-ordre).
Les anoures regroupent environ 3800 espèces. Ils sont caractérisés par :
- L’absence de queue chez les adultes.
- Les pattes postérieures sont longues, repliées en « Z » (pour le saut).
- Présence d’une palmure entre les doigts des mains et ceux des pieds.
- La larve est un têtard avec des branchies.
- Le nombre de vertèbres est réduit (5 à  .
.
- Radius / cubitus et tibia/péroné sont soudés
Ces anoures sont ensuite divisés en famille à partir de la structure des vertèbres.
a La famille des Bufonidés.
Il y a absence de dents sur les maxillaires. Exemple, le crapaud vulgaire ou Bufo bufo ; il vit en milieu terrestre. Il ne retourne à l’eau que pour la reproduction.
b La famille des Hylidés (ou rainettes).
Ces espèces sont arboricoles (dans les arbres). On peut observer une courbure des dernières phalanges qui donnent des crochets.
Exemple : Hyla arborea.
c La famille des Ranidés.
Ce sont toutes les espèces du type Rana.
2 Les urodèles.
Les urodèles regroupent environ 360 espèces.
Il y a une persistance de la queue chez les adultes ; la fécondation est semi-interne ; les branchies persistent jusqu’à la fin de la métamorphose ; ils n’ont pas de tympan.
On trouve plusieurs ordres comme les Salamandridae (salamandre et triton). Le plus grand des urodèles est Megalobatrachus japonicus (la salamandre géante) qui fait 1 mètre 60 pour 10 kilogrammes. Il appartient aux Cryptobronchidae.
3 Les apodes.
Les apodes sont serpentiformes. Ils mesurent entre 10 centimètres et un mètre. Ils ont des écailles dans le derme ; leurs yeux sont atrophiés et les adultes pondent dans des terriers proches de pièces d’eau.
Les larves atteignent un développement avancé à l’intérieur de l’œuf. L’éclosion donne des jeunes à respiration pulmonaire.
B La reproduction chez les amphibiens.
1 Les anoures.
Les mâles et les femelles vont se réunir dans des pièces d’eau où les mâles sont bruyant. Il y a deux types d’accouplement selon la position du mâle sur la femelle. L’accouplement axillaire et l’accouplement lombaire.
La fécondation est externe et 2000 à 6000 œufs sont pondus. Les œufs fécondés sont protégés par une enveloppe visqueuse pour éviter la dispersion. En plus de l’enveloppe, certaines espèces produisent une écume protectrice formée à partir de sécrétions cloacales, d’eau et de bulles d’air. Le tout entoure la ponte et la partie extérieure se durcie, ce qui permet aux œufs de résister à de courtes sécheresses.
De ces œufs vont éclore des têtards (à tête globuleuse). La tête possède un bec à dents cornées. Le reste du corps est allongé, avec un aplatissement latéral. Dorsalement et ventralement, des replis cutanés donnent des « nageoires ». Ces larves ont un régime alimentaire herbivore. Les têtards vivent 3 à 4 mois dans l’eau avant de subir la métamorphose et de devenir de jeunes grenouilles pouvant quitter le milieu aquatique.
Toutefois, toutes les espèces d’anoures ne se reproduisent pas dans l’eau. Certains pondent sur des rochers ou des plantes surplombant une pièce d’eau. A l’éclosion, les larves tombent dans l’eau.
2 Les urodèles.
Exemple du Triton palmé.
Ces tritons passent l’hiver le plus souvent dans l’eau. Au printemps, l’activité est importante, avec les font qui font une parade nuptiale (ils replient leur queue sur le coté et frétillent). Les mâles déposent un spermatophore (contenant des spermatozoïdes) dans l’eau et reculent. La femelle va suivre son mâle et passer sur le spermatophore. Elle va le capter avec ses lèvres cloacales ; il y a alors fécondation interne ou semi-interne.
Après la fécondation, la femelle pond ses œufs sur les feuilles de plantes aquatiques qu’elle va replier.
Les larves ont des branchies extérieures bien visibles et les membres sont beaucoup plus grêles que chez les adultes.
Ces larves ont une croissance rapide et la métamorphose a lieu en fin d’été.
C La métamorphose chez les amphibiens.
La métamorphose est un ensemble de processus qui aboutissent d’un plan de fonctionnement et d’organisation à un autre. Il y a des modifications structurales et fonctionnelles (apparition de nouveaux organes et/ou modifications et/ou disparition d’organes larvaires). Le déterminisme est de nature hormonale.
La métamorphose est plus complexe chez les anoures que chez les urodèles.
1 Les organes nouveaux.
Il y a apparition de vraies dents, de paupières, de la langue, de glandes cutanéespoumons et de membres (pour les anoures uniquement). pluricellulaires, de
Les pattes postérieures apparaissent en premier. Les pattes antérieures commencent à se développer dans les chambres branchiales.
2 Les transformations.
Il y a transformation de l’appareil circulatoire, de la musculature, de la peau. Il y a raccourcissement de l’intestin. La bouche s’agrandit et les yeux font saillie.
3 Les disparitions.
Le bec et les dents cornées disparaissent, ainsi que les branchies et la queue chez les anoures.
4 Les modifications de la métamorphose et de la fécondation.
Il peut y avoir réduction ou suppression de la métamorphose. Chez certains anoures tropicaux qui pondent des œufs riches en vitellus, il y a disparition du stade têtard. Les œufs donnent des jeunes semblables aux adultes. Le nombre d’œufs pondus est réduit à cause de la grande quantité de vitellus à produire.
De plus, tous les anoures et urodèles ne libèrent pas forcément les œufs dans le milieu extérieur. Par exemple, le crapaud accoucheur récupère les œufs fécondés (forment un long chapelet). Il les entortille autour de ses pattes postérieures. Il les garde pendant environ trois semaines. Quand les têtards sont près à sortit, le mâle va vers un point d’eau où les larves seront libérées.
Il existe d’autres espèces d’anoures qui portent les œufs sur le dos. La peau de ce dos se gonfle et sécrète une substance protectrice qui enveloppe l’ensemble de la ponte.
D’autres espèces portent les œufs et les larves au niveau de poches incubatrices (parfois même dans les sacs vocaux). On trouve dans ce cas les espèces arboricoles qui portent leurs œufs dans une poche dorsale.
Un espèce d’anoures australiens va, après la fécondation, avaler les œufs et les incuber dans l’estomac. Les femelles cessent alors de s’alimenter. La capsule des œufs synthétise une substance qui arrête la production d’enzymes digestives chez la mère. A l’éclosion, les têtards sont expulsés par la bouche. La femelle pourra alors recommencer à se nourrir.
On trouve aussi des espèces vivipares. Chez elles, il existe au niveau de l’oviducte, une poche incubatrice dans laquelle les têtards ont des rapports nourriciers avec la mère. Ce phénomène est comparable à ce qu’il se passe chez les mammifères.
Chapitre 7 : L’Hominisation.
Généralités.
L’hominisation, en biologie générale, peut être vue d’un point de vue du vivant et c’est alors une étude d’organismes qui vont se multiplier et évoluer au fil du temps, ou bien, du point de vue des pressions écologiques et ce qui pousse l’Homme à évoluer.
L’Homme n’est classé dans le règne animal que depuis le XVIIIème siècle.
Au cours de l’évolution, les facteurs écologiques ont été remplacés par des facteurs culturels (de plus en plus) : l’Homme s’est affranchi des facteurs écologiques et a conquis tous les milieux. Par exemple, pour sa thermorégulation, quand l’Homme a froid, il met un pull au lieu d’attendre l’augmentation de sa thermogenèse.
Quatre grandes étapes dans l’évolution humaine.
I : L’origine de la lignée humaine.
Cette origine est située entre 6 et 2 millions d’années. C’est l’apparition de fossiles qui progressivement mènent au genre Homo.
II : Origine de l’Homme moderne (Homo sapiens).
Quand est apparu l’Homme. Quelles ont été les modalités de l’apparition de cette espèce (entre 500 000 et 100 000 ans).
Les premiers individus de cette espèce étaient des chasseurs-cueilleurs. Ils étaient peu nombreux (faible densité de population).
III : La néolithisation.
La néolithisation est le passage d’une économie de subsistance de chasseurs-cueilleurs à une économie d’agriculture (plus forte densité). C’est l’apparition d’une forte démographie.
IV : La transition démographique.
En Europe, cette transition démographique à eu lieu à la fin du XVIIIème siècle avec l’augmentation des conditions de vie et l’apparition des vaccins (par Jenner). Il y a alors eu un « boum » démographique. Par la suite, il y a eu la découverte des antibiotiques (XXème siècle) et la découverte de la pilule, moyen de contraception réellement efficace.
I L’Hominisation.
L’hominisation est l’apparition d’hominidés bipèdes ; l’apparition de la lignée conduisant au genre Homo.
Actuellement, l’Homme est le seul bipède. Il y a eu une époque, entre 4 et 2 millions d’années, où on avait plusieurs familles d’animaux bipèdes.
A partir de quand parle-t-on du genre humain ?
Les animaux les plus proches de nous sont les grands singes : gorilles, chimpanzés (dont le Bonobo) et orang-outang. Ces animaux sont proches sur les plans anatomique, physiologique et génétique.
Sur le plan anatomique, il existe des différences caractéristiques entre les singes et l’Homme :
- Pour la bipédie, le squelette des membres inférieurs (et surtout le fémur) est modifié mais aussi le bassin, la colonne (tronc) et le trou occipital. Chez l’Homme le fémur possède une orientation particulière.
- Les différences au niveau du crâne.
· Le neurocrâne : il est déterminé par le cerveau. La capacité crânienne humaine est d’environ 1440cm3, avec des variations comprises entre 1000 et 2000 cm3.
Pour les singes (chimpanzés), la capacité moyenne du neurocrâne est d’environ 300 cm3. Certains cerveaux de gorille atteignent un volume de 440cm3.
La capacité crânienne mise en place chez l’Homme au cours de l’évolution est telle que l’Homme est la seule espèce à avoir des problèmes lors de la mise bas.
· Le splanchnocrâne : chez l’Homme, le système masticateur est réduit, ce qui se répercute sur les dents. La forme des dents est particulière.
Chez les singes, la mastication est réalisée par un système musculaire ultra-puissant : il y a alors un développement de super-structures osseuses permettant l’absorption des chocs ce sont les bourrelets sus-orbitaires.
D’un point de vue non anatomique, la différence principale de l’Homme est la conscience de soi. Mais pour ce qui est de la retrouver dans les fossiles… Toutefois, il existe les outils et leur utilisation. Pour caractériser une espèce ayant conscience de soi, on essaie de relever les traces d’outils complexes nécessitant l’utilisation de chaînes opératoires complexes. Les premiers outils complexes ont plus de 2,5 millions d’années : ce sont des blocs avec des éclats de pierre. Une fois que l’on a les outils, il faut retrouver l’animal capable de faire ces actions.
On a pensé à l’australopithèque (que l’on croyait humain) mais son volume crânien n’était que de 330cm3 et ne devait donc pas avoir de cerveau complexe, soit l’ancêtre Homo est non connu.
A 6 millions d’années, on a retrouvé un fossile dont on n’a pas la capacité crânienne mais dont on a le fémur et quelques dents : « Millenium ancestor ». Ce fémur indique la bipédie et les dents indique que le système masticateur était déjà réduit. Ce serait ces animaux qui auraient eu des descendants ayant fait ces outils.
Avant les six derniers millions d’années, on a le problème de l’absence de gisement de fossiles.
A partir de quand parler du genre Homo ?
A 2 millions d’années, on a trouvé des sujets (en Afrique) qui atteignaient 600cm3 de neurocrâne ; ils ont été classés dans le genre Homo. A partir de 1,8 millions d’années, on trouve des fossiles avec 700cm3 de neurocrâne et avec un système masticateur réduit. L’ensemble des fossiles que l’on trouve de cette époque sont tous d’Afrique.
Quels ont pu être les facteurs écologiques ayant permis l’arrivée du genre Homo ?
Une théorie controversée : Entre 10 et 2 millions d’années, il y a eu une sécheresse en Afrique : toute la partie Est serait devenue une savane. Les déplacements se seraient fait de plus en plus au sol, d’où, la bipédie. La partie Ouest n’aurait pas subit de changement ; les espèces n’auraient pas évolué.
Après deux millions d’années, à partir de 1,8 millions d’années, on a des restes d’hominidés en Eurasie ce sont les Homo erectus qui vont conquérir l’Asie. L’Amérique et l’Australie ont été atteinte récemment : 20 000 ans pour l’Amérique et 60 000 ans pour l’Australie.
A 500 000 ans, on a des fossiles à plus de 1000 cm3 de neurocrâne. On peut observer des restes d’innovations culturelles ; de plus, le feu a été domestiqué.
II L’origine de l’Homme moderne.
Plusieurs théories s’affrontent quant à l’origine de l’Homme moderne.
Théorie 1 : La théorie polycentrique.
Entre 500 000 ans et 60 000, en Afrique et en Asie, on trouve des fossiles d’Homo sapiens sapiens. Il doit y avoir une évolution sur place des Homo erectus Africains et des Homo erectus asiatiques, afin de donner les populations actuelles. C’est une évolution graduelle de sujets engagés graduellement dans la lignée d’Homo sapiens.
Cette hypothèse est basée sur des fossiles et leurs particularités anatomiques. L’Homo erectus asiatique présente des caractéristiques fréquentes : les incisives supérieures sont mises en pelle, les premières molaires inférieures ont trois racines, l’os occipital a un point supérieur d’ossification os triangulaire aussi connu sous le nom « d’os des Incas ».
Théorie 2 : La théorie monocentrique.
Cette théorie est défendue par les généticiens.
Quand on voit la variabilité génétique des populations modernes, on voit que celle-ci est faible : donc, au maximum, notre ancêtre commun date d’il y a 500 000 à 100 000 ans. l’humanité viendrait d’une seule souche apparue dans cet intervalle de temps. Or, actuellement, c’est en Afrique que l’on a le maximum de variabilité : ce n’est plus une naissance graduelle mais une naissance par clade. Une mutation d’Homo erectus africain aurait donné cet ancêtre qui aurait reconquis la planète. Toute
Toutefois, on peut se demander pourquoi Homo erectus a disparu.
Les H. erectus devaient avoir un proto-langage alors que H. sapiens ont un langage.
Les généticiens travaillent sur les populations actuelles et font des études de variabilité. Ils se basent sur des marqueurs comme les groupes sanguins mais aussi les marqueurs nucléaires non liés aux chromosomes sexuels (ils sont peu utilisés), les marqueurs liés au chromosome Y (ils sont intéressants) car ils permettent de remonter dans une lignée et les marqueurs d’ADN mitochondrial (transmis par la mère).
Il va y avoir des études (statistiques) de la diversité dans une même population et entre différentes populations. Il faut prendre en compte toutes les mutations, ce qui fait apparaître le problème de l’horloge moléculaire. Beaucoup d’hypothèses ont été formulées. Pour s’aider dans l’horloge moléculaire, les généticiens ont pris le cas de l’Australie, colonisée il y a 60 000 ans. Les aborigènes ont été séparés des autres populations pendant des dizaines de millier d’année. Ils ont donc subit des mutations sans mélanger leur génome avec d’autres populations. En connaissant le nombre de mutations et la durée nécessaire à celles-ci, on peut avoir une idée de l’horloge moléculaire.
La variabilité de l’ADN mitochondrial et du chromosome Y est relativement faible et quand on applique l’horloge moléculaire, on obtient l’ancienneté des variations. Elles sont comprises entre 150 000 et 200 000 ans pour l’ADN mitochondrial et comprises entre 60 000 et 100 000 ans pour le chromosome Y.
Les populations présentant une variabilité maximale sont d’Afrique de l’Est (Ethiopie, Somalie). Pour les généticiens, il n’y a qu’une origine de l’Homme moderne, car il n’a pas assez d’ancienneté.
De plus, la possibilité d’avoir deux apparitions d’une même espèce dans des lieux différents dans le monde est quasiment nulle. Les plus anciens fossiles connus de l’Homme moderne sont en Afrique et date de la période donnée par les généticiens.
Le décalage existant entre les variations données par l’ADN mitochondrial et le chromosome Y peuvent être expliquées du fait que les vitesses de variation de certains marqueurs peuvent changer au cours du temps. De plus, on n’est pas sur de prendre le bon marqueur ni la bonne mutation. Donc, ce que l’on voit dépend du marqueur que l’on regarde.
On peut aussi se demander pourquoi cette espèce moderne aurait dominé et remplacé les autres : elle avait le langage (le langage est différent de communication) qui est associé à des possibilités d’abstraction.
Théorie 3 : Théorie de l’hypothèse réticulée.
Cette théorie tient compte des données de paléontologie et de génétique. Elle est basée sur le fait que la définition d’espèce est donnée par des scientifiques et que dans la nature, il y a de nombreuses exceptions.
L’Homme moderne est apparu il y a entre 100 et 200 000 ans, et, grâce à ses capacités, il aurait supplanté les autres populations ou se serait croisé avec elles. Ceci expliquerait l’existence de certains marqueurs morphologiques.
III Le cas du peuplement de l’Europe et des Néandertaliens.
L’Europe est un schéma à part. Il y a un gisement archéologique de 1,2 million d’années mais sans ossements humain (qu’avec des objets). Vers 500 000 ans, on a quelques restes humains, attribuables à des Homo erectus. A partir de 200 000 ans et surtout 100 000 ans, les Homo erectus évoluent et vont se différencier de plus en plus : il y a apparition de caractéristiques différenciées. Ces nouveaux humanoïdes portent des caractères archaïquestorus (bourrelet frontal) ; des caractères partagés avec l’Homme moderne (la capacité crânienne de ces humanoïdes est supérieure ou égale à celle des sapiens) ; des caractères morphologiques propres, comme un mode de croissance du crâne qui est à l’origine d’un crâne très allongé : « face en museau ». comme un
A 70 000 ans, toute la population de l’Europe porte ces caractères.
On hésite encore à mettre cette population comme une espèce à part (Homo neandertalansis) ou comme une sous-espèce (Homo sapiens neandertalansis).
Ce groupe, entre 40 000 et 20 000 ans avant JC, va disparaître. Il est associé en Europe à une industrie en pierre (Moustérienne), faite d’éclats. La disparition est brusque en certains endroits, mais il existe encore quelques lieux où cette «espèce» persiste. En même temps, il va se mettre en place un nouveau type d’industrie de pierre en lame(l’Aurignacien), associée à des squelettes d’Hommes modernes.
Il semble que ce soit l’Homme moderne qui ait effectué les premières peintures.
· Il y aurait eu une différenciation des Homo erectus car l’Europe, à ce moment là, était un cul de sac : au cours des glaciations quaternaires, les glaciers ont fait un arc de cercle de la Belgique à la mer noire et cette population s’est retrouvée isolée. Il y a eu l’isolation mais aussi la faible densité de population. Il semblerait y avoir eu un phénomène comparable en Indonésie.
· On voit que l’Homme moderne a reconquis l’Europe à partir de 40 000 ans. Il y a ensuite une diffusion qui dure 20 000 ans. Les modalités de remplacement sont de différents types :
- Remplacement sans croisement car les deux populations sont différentes. Il peut y avoir une barrière entre les deux espèces.
- Les néandertaliens ont la même capacité crânienne que les Homo sapiens ; on peut penser qu’ils avaient aussi une pensée symbolique car ils enterraient leurs morts. Quand il y a eu contact entre les deux espèces, il y aurait eu évolution de la technologie moustérienne. Sur le plan anatomique, on peut voir que les néandertaliens avaient la possibilité de parler car ils possèdent l’os hyoïde, comme les sapiens.
Il a été fait des études sur les fossiles pour savoir si l’on a un hybride, mais l’interprétation dans ces cas là est trop subjective.
Il a aussi été réalisé des études d’ADN néandertaliens faites à partir d’ADN mitochondrial extrait d’os à partir de trois sujets. Ces études ont bien confirmé que les néandertaliens s’étaient séparés il y a 500 000 ans. De plus, la variabilité de cet ADN est faible et ces variations ne recoupent quasiment pas celles des populations actuelles. Donc, soit personne ne dérive des néandertaliens (barrière entre espèces), soit il y a eu quelques croisements (donc pas de barrière) qui n’ont laissés que de rares traces.
IV De qui descendons nous ?
On considère que les premiers Homo sapiens étaient tous chasseurs-cueilleurs. A la suite de la disparition des glaciers, on a eu en différents points du globe, l’apparition d’un nouveau mode de subsistance : l’agriculture. Celle-ci est apparue dans les foyers primaires de néolithisation. Dans ces foyers, il existaient des plantes sauvages qui ont été domestiquées.
Le premier foyer européen est apparu dans le proche orient, avec des blés et des légumineuses sauvages. Progressivement, la population s’est accrue et a commencé à diffuser (elle a été suivie par les foyers écologiques). Des populations issues de ce foyer primaire se sont dirigées vers chypre puis vers les balkans. De là, elles ont pris deux voies différentes : une voie danubienne qui est arrivée dans le bassin parisien vers –4500 avant JC et une voie qui a suivi la côte méditerranéenne et qui est arrivée vers –5500 avant JC.
Ces néolithiques avaient une démographie beaucoup plus importante que les chasseurs cueilleurs. Ils les ont remplacés ou convertis à l’agriculture en leur faisant prendre la même démographie.
Avant, les discussions sur ces populations étaient basées sur des données archéologiques. Se sont ensuite rajoutées des données génétiques, sur le langage et enfin, des données apportées par le chromosome Y et l’ADN mitochondrial.
On peut voir que la quasi-totalité des langues européennes actuelles sont parentes entre elles : ce sont des langues indo-européennes. Une hypothèse est que le noyau de cette langue était situé en Turquie et que celle-ci se serait déplacée d’Ouest en est ; ce qui semble correct.
Le problème du Basque.
Le Basque n’a aucun équivalent connu. Une hypothèse est que cette langue pourrait avoir comme origine les chasseurs cueilleurs. Il y aurait eu un replis de cette population dans la partie Ouest des pyrénées. On peut aussi voir que les Basques ont un profil génétique particulier, comme le groupe sanguin O rhésus -, très présent dans cette population.
Aujourd’hui, on considère que moins de 20% des gènes que nous possédons proviendrait des chasseurs-cueilleurs. On trouverait un maximum de ces gènes vers l’Ouest des Pyrénées, avec un gradient qui diminue jusqu’à la Garonne :
- Soit, il y a eu un tout petit noyau avec des personnes qui auraient diffusées ;
- Soit, les agriculteurs auraient repoussé cette population vers la partie Ouest des Pyrénées.