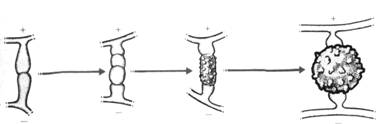
- monoïque si les gamètes mâles et femelles sont sur le même individu
- dioïque si les deux types de gamètes sont sur des individus différents.
Chapitre 1 : LES BACTERIES.
On a deux grands groupes, les archéobactéries et les eubactéries.
La classification est basée sur des caractères génotypiques, sur la filiation évolutive (techniques de séquençage, types de parois, …). Par exemple, la paroi des archéobactéries ne renferme pas d’acide muramique qui est le composant typique des peptidoglycanes. Les lipides membranaires présentent des chaînes aliphatiques ramifiées, alors que chez les eubactéries, ces mêmes chaînes ne le sont pas. De plus, les archéobactéries possèdent des gènes discontinus.
I Les eubactéries.
A Morphologie des eubactéries.
Elles sont en général unicellulaires mais on les trouve parfois sous forme de colonies plus ou moins structurées. Les cellules sont soit sphériques : coccoïdes, soit en bâtonnets : bacillus.
L’enveloppe : c’est la membrane la plus interne, elle est cytoplasmique, rarement en contact avec le milieu externe dont elle est séparée par la paroi. Chez certaines espèces, la membrane cytoplasmique peut s’invaginer pour donner un empilement de membrane interne : le mésosome. Cela se retrouve particulièrement chez les bactéries photosynthétiques ou, chez les bactéries ayant des échanges gazeux importants.
La coloration de Gram permet de séparer les eubactéries en deux groupes : les g+ et les g-. Cette coloration est révélatrice d’une différence de structure de la paroi.
- Chez les g+, la membrane cytoplasmique est recouverte d’une zone épaisse de peptidoglycane qui enferme la cellule dans une coque relativement rigide et épaisse. Cette gangue est responsable de la forme cellulaire et est chargée de constituants secondaires comme l’acide teïchoïque.
- Chez les g-, la couche de peptidoglycanes est faible et est recouverte d’une seconde membrane externe de composition spéciale renfermant des LippoPolySaccharides (LPS). Entre la membrane interne et la membrane externe, se situe un espace étroit appelé le Périplasme où se situe le peptidoglycane.
Les bactéries s’entourent souvent d’une enveloppe supplémentaire plus ou moins structurée, parfois épaisse, appelée le Glycocalyx.
B Le génome.
Dans une bactérie, il est représenté par un double brin d’ADN circulaire. Le génome est parfois complété par des anneaux d’ADN supplémentaires plus petits : ce sont les plasmides. Ils offrent au monde bactérien des possibilités extraordinaires d’adaptation car ils sont souvent transmissibles d’une cellule à l’autre. Les bactéries échangent entre elles des portions de séquences d’ADN ou de plasmides qui peuvent s’intégrer complètement dans le génome de la bactérie receveuse.
C Les cils et les flagelles.
Ils sont comparables aux flagelles des cellules mobiles des algues. Ils sont constitués par un petit nombre de fibrilles et s’insèrent au niveau du plasmalème sur un blépharoblaste. La protéine constituant ces flagelles est élastique et contractile comme la myosine des muscles.
D La multiplication asexuée.
Les bactéries se divisent par scissiparité. La division cellulaire peut-être rapide (de 20 à 30 minutes), à partir d’une cellule, on peut en obtenir jusqu’à 10^9 en 24 heures. C’est un phénomène de parasexualité. Chez certaines bactéries, il existe des processus parasexués aboutissant à des recombinaisons génétiques voisines de celles résultant de la reproduction sexuée des eucaryotes.
On connaît trois processus : transformation, transduction, conjugaison qui ont des caractéristiques générales qui permettent de les distinguer de la reproduction sexuée des eucaryotes.
Chez ces derniers, la contribution des deux gamètes pour constituer le matériel génétique du zygote est symétrique. Un zygote diploïde redevient haploïde à la suite d’une méiose. Dans les processus parasexués, il n’y a pas de gamètes, mais deux cellules à rôle opposé : un parent donneur qui introduit dans l’autre parent (receveur ou accepteur), une partie de son matériel génétique et donne un mérozygote qui contient la totalité du matériel génétique de l’accepteur. Ce matériel génétique est appelé endogénote.
De plus, un ou plusieurs fragments d’ADN du donneur (exogénote) rentre dans ce nouveau matériel génétique mais les exogénotes sont incapables de se multiplier et finissent par disparaître. De temps en temps, l’exogénote peut s’intercaler avec la région homologue de l’ADN du receveur. Par la suite, des descendances du mérozygote vont apparaître et donneront des bactéries à matériel génétique recombiné. Toutefois, la majeure partie du génome provient du receveur.
1 La transformation.
Exemple avec Streptococcus pneumoniae : Un échange génétique peut se produire, quand, dans un milieu de culture où se développe une population bactérienne avec un génotype donné, on introduit de l’ADN correspondant à un génotype différent. La transformation ne peut se produire que si les bactéries sont dans un état «compétent ».
Dans le cas de l’agent de la pneumonie (g+), l’état de compétence est conféré par un facteur de compétence (une protéine soluble de faible poids moléculaire, produite et excrétée par les bactéries dans le milieu de culture) qui est absorbé sur certains sites de la surface cellulaire. Ce facteur induit une cascade de réactions permettant l’entrée dans la cellule de l’ADN transformant.
Si l’ADN est originaire d’un organisme différent mais s’il présente suffisamment d’homologies avec celui de l’endogénote, il est intégré à ce dernier sous forme de courtes séquences. La bactérie réceptrice, en intégrant une partie de l’information génétique de l’ADN donneur, acquière donc, une ou plusieurs caractéristiques du donneur.
2 La transduction.
Ce mécanisme de recombinaison génétique s’opère par l’intermédiaire d’un bactériophage. Il y a infection d’une bactérie par un virus (virion), qui se multiplie à l’intérieur et qui provoque sa destruction. Pendant cette multiplication, ce virus peut intégrer à son ADN une partie du génome de la cellule infectée. Au cours de l’infection d’une nouvelle bactérie, le virion introduit dans celle-ci, une séquence du génome de la cellule infectée précédemment. Ces virions sont appelés : « vecteur de transduction ».
3 La conjugaison (sur Escherichia coli).
Une véritable différenciation de type sexuel existe. Deux bactéries entrent en contact par un pont cytoplasmique. La bactérie mâle est plus petite que la bactérie femelle (réceptrice). La bactérie mâle injecte un brin d’ADN représenté par un plasmide ou un chromosome.
Un trait dominant du monde bactérien est une grande variété et un important métabolisme. Les bactéries peuvent fixer l’azote atmosphérique.
Exemple : Des bactéries sont à l’origine du gaz naturel et de la houille. Elles ont donc un rôle fondamental. Sur l’Homme, elles peuvent avoir un rôle bénéfique ou pathogène.
Les bactéries peuvent être :
- aérobies strictes.
- aérobies facultatives (elles vivent avec ou sans oxygène).
- anaérobies strictes (elles vivent sans oxygène).
- anaérobies qui supportent la présence d'oxygène en faibles concentrations.
Les bactéries anaérobies sont les plus anciennes. Certaines bactéries, comme les végétaux, sont capables d’utiliser les radiations lumineuses comme source d’énergie. Ce sont des phototrophes, mais leurs pigments sont différents de ceux des végétaux. En général, la photosynthèse a lieu en milieu, à peu près, anaérobie.
Les bactéries qui effectuent toutes leurs synthèses à partir du CO2 comme seule source de carbone sont dites «photoautotrophes ».
D’autres bactéries vivent au dépend des composés organiques tout en continuant à utiliser l’énergie lumineuse. Ce sont les «photohétérotrophes ».
Des bactéries vertes utilisent le CO2 comme source de carbone et H2S comme source de pouvoir réducteur. Ces bactéries sont dites : « chimio-litho-hétérotrophes ».
 Les bactéries peuvent vivre partout.
Les bactéries peuvent vivre partout.
Remarque : Les pigments permettant la photosynthèse sont les bactériophylles et les caroténoïdes.
II Les cyanobactéries.
C’est le deuxième grand groupe des procaryotes. On les appelle aussi cyanophycées ou blue-green algae.
A Généralités.
Les cyanobactéries n’ont pas de recombinaison génétique. Au niveau des pigments, on note la présence de chlorophylle A, ainsi que d’autres pigments : les billiprotéines qui sont solubles dans l’eau (exemple : la Phycoérythrine qui est rouge et la phycocyanine qui est bleue).
Au microscope, la cellule bactérienne apparaît souvent homogène car elle n’a pas de plastes individualisés. Toutefois, on arrive à distinguer une zone périphérique : le chromatoplasme, et une partie centrale : le centroplasme ou nucléoplasme. La cellule est entourée d’une paroi épaisse. Celle-ci est similaire à la paroi des gram-. Dans un certain nombre de genres, on note la présence d’une gaine. Celle-ci peut avoir une structure lamellaire très épaisse. Les cellules sont dans une sorte de gelée.
B Structure et fonctions.
1Le chromatoplasma.
C’est une partie très colorée par les pigments. On y trouve des thylacoïdes (se sont des sacs aplatis sur lesquels est fixée la chlorophylle). Ils sont dispersés dans le centroplasme, mais ne sont jamais entouré de parois pour former des chloroplastes.
Le complexe pigmentaire. Il est constitué par la chlorophylle A qui est fixée sur les thylacoïdes, et par les caroténoïdes.
L’énergie lumineuse que peuvent capter les caroténoïdes est faible mais ils peuvent avoir un rôle écologique important car ils protègent contre une trop forte intensité lumineuse et empêchent donc la photo-inhibition.
Les billiprotéines sont des pigments, excellents capteurs de l’énergie lumineuse et la retransmettent presque à 100% à la chlorophylle. Ces billiprotéines donnent un avantage car la présence de ces deux pigments permet de capter tout le spectre de la lumière (entre 400 et 800 nm). Quand le milieu est carencé en azote, les cyanobactéries utilisent celui contenu dans les billiprotéines.
2 Les inclusions cellulaires.
Les granules de carboxysome : elles sont le lieu de localisation des enzymes fixant le CO2 (Rubisco)
Les granules de polyphosphates (= la volutine). Ils sont métachromatiques (s’ils sont colorés par le bleu de méthylène ils seront rouges). Ils sont le lieu d’accumulation du phosphate. Ils sont utilisés quand le milieu extérieur est carencé en phosphate.
Les granules de cyanophycine. Ce sont des réserves d’azote qui peuvent être utilisées des deux côtés de la paroi cellulaire.
Ces granules se constituent quand le milieu est riche en un élément. C’est un avantage dans la compétition entre cellules. A partir de ses réserves, une cellule peut se multiplier 7 à 8 fois.
Chez les cyanobactéries planctoniques, on trouve très souvent des pseudo-vacuoles gazeuses (ce sont de petits cylindres creux, remplis d’air, leur diamètre est à peu près de 70 nm). Leur paroi est perméable à l’eau : c’est un espace creux en équilibre avec les gaz dissous dans le cytoplasme
Leur rôle est de permettre aux cyanobactéries de faire des migrations verticales soient journalières, soient, plus étalées dans le temps. Quand ces bactéries sont soumises à une trop faible intensité lumineuse, il y a multiplication des pseudo-vacuoles gazeuse : ce qui permet une remontée vers la surface.
è Donc : plus l’intensité lumineuse augmente, plus la pression osmotique augmente. On assiste donc a un collapsus ou dégonflement de ces vacuoles qui entraîne la descente des ces bactéries.
Le génome : Il est constitué de fibrilles d’ADN localisées dans le nucléoplasma.
3 Cyanobactéries coloniales et multiplication asexuée.
Certaines cyanobactéries (coloniales) possèdent des cellules spéciales appelées «hétérocystes ». Ces cellules se distinguent par une couleur plus verdâtre, moins dense, avec une paroi plus épaisse et surtout avec à chaque extrémité, la présence d’un pore qui la met en contact avec les cellules contiguës du filament. Les hétérocystes perdent leur carboxysomes (et l’enzyme Rubisco [Ribulose 1,5 diphosphate carboxylase] qui fixe le CO2), et ne peuvent donc plus effectuer la photosynthèse, et ne peuvent en conséquence, plus rejeter d’oxygène.
Les hétérocystes sont considérés comme les cellules les plus aptes à fixer l’azote atmosphérique. Tout autour des hétérocystes, il n’y a pas d’oxygène, c’est ce qui permet un meilleur fonctionnement de la nitrogénase (c’est l’enzyme qui fixe l’azote).
En milieu anaérobie, toutes les cellules fixent l’azote atmosphérique. Les hétérocystes ont la faculté de produire des askinètes. Ce sont des spores de résistances : c’est une cellule normale qui grandit, se remplie de matière organique et se protège avec une épaisse membrane, puis se laisse tomber au fond du milieu, puis remontera plus tard pour recoloniser le milieu.
Remarque : il existe de vrais et de fausses ramification chez les cyanobactéries.
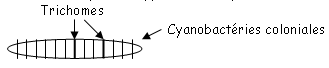
La multiplication asexuée : certaines formes de cyanobactéries forment des endospores ou nannocystes, d’autres forment des exospores. Beaucoup de cyanobactéries se multiplient grâce à des spores pluricellulaires qui sont appelés hormospores.
Ecologie des cyanobactéries :
Elles sont rencontrées dans tous les milieux. Dans certains cas, elles ont un rôle utile, par exemple, dans certains cours d’eau, elles fixent l’azote et servent ainsi d’engrais naturel. Elles peuvent aussi avoir des effets négatifs : elles peuvent sécréter des toxines qui seront toxiques pour les autres habitants du milieu. Elles peuvent aussi être néfastes par leur nombre.
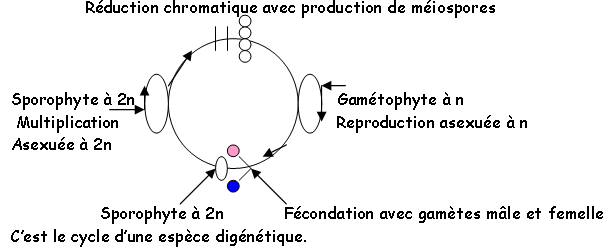
Chapitre 2 : Sexualité et compatibilité.
Si la reproduction sexuée suppose la formation de gamètes des deux sexes, elle exige en outre la compatibilité : les gamètes mâles et femelles doivent avoir le pouvoir de s’unir.
I La différenciation sexuelle.
Elle est génotypique ; si à la méiose, une ségrégation est responsable de la formation en nombre égal de deux catégories de méiospores, on a alors 50% de mâles et 50% de femelles.
A contrario, s’il n’y a pas de ségrégation sexuelle méiotique, la manifestation de la sexualité sera beaucoup plus tardive et seulement d’ordre phénotypique. Cette différenciation s’établit sous l’influence de facteurs internes et externes. Ces deux possibilités se rencontrent chez les algues et chez les mycètes.
Cette présentation du déterminisme de la sexualité est simplifiée car il ne tient pas compte du phénomène de sexualité relative.
On a 4 types de gènes : masculinisant fort, masculinisant faible, féminisant et neutre, qui sont responsables de 4 catégories de thalles : Mâle fort, mâle faible, femelle forte, femelle faible. Si un gamète mâle faible se comporte comme un mâle avec les gamètes femelles (forte et faible), il peut se comporter comme un gamète femelle avec un mâle fort.
Quand les gamètes mâles et femelles arrivent à maturité en même temps sur un même thalle, celui-ci appartient à une espèce monoïque, ce qui n’implique pas forcément de reproduction monoïque (autofertilité ou autostérilité).
Si l’on a plusieurs thalles, on observe une interfertilité ou une interstérilité selon les cas. Pour les dioïques, le voisinage d’un thalle mâle et d’un thalle femelle mature, n’est pas suffisant pour qu’il y ait fécondation (il y a stérilité ou fertilité).
II Homothallisme et hétérothallisme.
A Homothallisme.
S’il y a incompatibilité, il existe, dans une espèce homothallique des thalles génétiquement différents.
Si l’espèce est monoïque, chaque thalle est autofertile mais l’interfertilité est possible avec les autres thalles.
Si l’espèce est dioïque, il faut deux thalles différents. Dans ce cas, tous les thalles mâles et femelles sont interfertiles.
Dans le cas des homothalliques, la recombinaison génétique est, sauf mutation, pratiquement nulle ou très faible pour les espèces monoïques. Si, chez des monoïques, les gamétocystes des deux sexes ne parviennent pas à maturité en même temps, l’autofertilité n’est plus que théorique, et l’interfertilité autorise dans ce cas, une certaine recombinaison génétique.
L’homothallisme se rencontre chez les algues et les mycètes. Toutefois les mycètes présentent plus souvent l’hétérothallisme.
B L’hétérothallisme.
On a un ou deux couples d’allèles qui sont responsables de la compatibilité et qui sont séparés pendant la méiose. Ce phénomène entraîne l’existence de deux catégories de thalles chez les monoïques (on parle d’hétérothallisme bipolaire)
Ces mêmes couples d’allèles entraînent l’existence de 4 catégories de thalles chez les dioïques (on parle d’hétérothallisme tétrapolaire).
1 Hétérothallisme bipolaire.
Une espèce monoïque est représentée par deux sortes de thalles. Les uns donnent le type conjugal plus (+), les autres donnent le type conjugal moins (-). La fécondation ne peut se faire qu’entre gamètes complémentaires + et -. L’autostérilité est donc obligatoire.
2 Hétérothallisme tétrapolaire.
Chez les dioïques, on a quatre catégories de thalles : les mâles +, les mâles -, les femelles +, les femelles -. L’interfertilité n’est réalisée qu’entre thalles de sexe et de type conjugaux différents. Si l'hétérothallisme tétrapolaire donne quatre types de thalles (A1B1, A1B2, A2B1, A2B2), on a interstérilité quand les thalles ne sont pas entièrement complémentaires et infertilités quand ils le sont.
La fécondation unie toujours les 4 allèles différents.
Chapitre 3 : Les Algues.
I Généralités.
Les algues sont des thallophytes. Elles se reproduisent grâce à des gamètes qui se forment dans les gamétocystes et se multiplient végétativement par des spores formées dans les sporocystes.
Les algues se caractérisent par la présence d’un noyau et de plastes, et par la reproduction sexuée.
En 1975, des chercheurs ont trouvé un procaryote : l’Olochron, qui vit en symbiose avec l’Aclidie et qui contient de la chlorophylle A et B. Il serait à l’origine de tous les végétaux supérieurs contenant ces deux types de chlorophylles.
Les algues existent dans tous les milieux humides, dans l’air. Dans les régions tropicales on les trouve même sur les murs des bâtiments. Elles peuvent être endophytes de certains protozoaires ou métazoaires. Certaines algues s’associent à des champignons et forment les lichens.
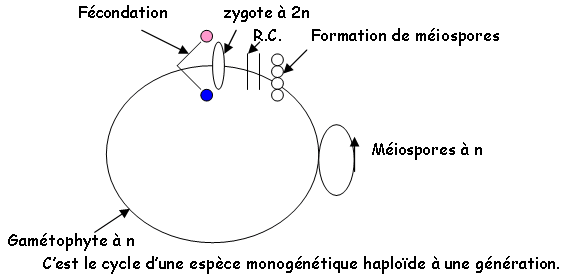
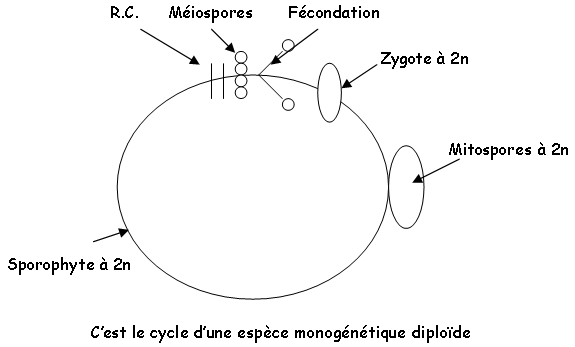
Au point de vue reproduction, on trouve trois cycles différents chez les algues : haplophasique, haplodiplophasique ou diplophasique. Elles ont des modes de fécondation divers : planogamie (gamètes flagellés), oogamie (gamète mâle mobile et femelle immobile), cystogamie (pas de gamète flagellé), trichogamie…
II Caractères morphologiques et cytologique des algues.
- Elles peuvent être unicellulaires, filamenteuses, ou parenchymateuses. Leur taille est très variable : de 3µm à 60m.
- Cytologie des algues : Le plastidum est appareil cinétique qui concerne les flagelles.
- Chez les algues, les plastes sont très variés et différents par leur morphologie, leur nombre, leurs dimensions, la structure et la composition chimique.
- La morphologie des plastes varie beaucoup d’un groupe à l’autre. Les algues considérées comme les moins évoluées ne contiennent qu’un plaste par cellule. Ce sont les archéons.
D’autres algues ont leurs plastes réunis en réseau par un tractus incolore : on dit que ces plastes sont mesplastidiés. Les formes les plus évoluées d’algues renferment de nombreux plastes indépendants, comme ceux des plantes supérieures qui sont dits néoplastidiés. Cette fragmentation successive des plastes entraîne une augmentation de la surface active par rapport au volume des substances plastidioles renfermées dans la cellule. Certaines formes évoluées ont une structure hétéroplastidiées. On assiste à une division du travail entre deux séries de plastes : des chloroplastes spécifiques de la photosynthèse et des leucoplastes ou amiloplastes spécifiques de l’élaboration et de l’accumulation d’amidon.
Remarque : Quel que soit leur nombre, les chloroplastes montrent un phototachisme très net offrant leur plus grande surface aux rayons lumineux suivant l’intensité de ceux-ci.
III Structure.
A L’enveloppe.
Le nombre de chloroplastes est témoin de l’origine de ces groupes. Chez les algues, on a par exemple :
- Les rhodophylles ont un chloroplaste entouré de deux membranes. En fait, ces eux membranes ont une origine différente
- Euglénophylles et dinophycées ont trois membranes autour des plastes. Pour les dinophycées, cela viendrait de trois symbioses successives et pour les euglénophylles, ce serait une cellule eucaryote qui aurait englobé une association déjà symbiotique avec des chloproplastes à deux membranes et peut-être quelques restes d’algues vertes.
- Dans le cas des organites à quatre membranes, il y aurait deus symbioses successives, une symbiose entre un ancêtre eucaryote et une cyanobactérie. Ce qui donnerait l’archétype d’une algue rouge qui ensuite aurait subit une symbiose avec un eucaryote flagellé incolore.
En microscopie électronique, on a mis en évidence l’existence d’un organisme nucléomorphe (vestige de noyau).
B Les chloroplastes.
Ce sont les plastes chlorophylliens des algues. Ils possèdent une structure lamellaire due aux thylacoïdes qui sont des sacs membraneux aplatis, au niveau desquels sont localisés les pigments des cyanobactéries. Ils ne sont pas entourés de membrane dans le cytoplasme.
Chez les algues rouges, les thylacoïdes sont séparés les une des autres. Les phycobilisomes sont à l’extérieur des thylacoïdes comme chez les cyanobactéries.
Chez les algues brunes (les phéophytes) diatomées et les chrysophycées, les thylacoïdes sont groupés par trois et accolés sur une grande surface. Chez les cryptophycées, les thylacoïdes sont groupés par paires.
Chez les algues vertes (A+B), les thylacoïdes forment un empilement irrégulier rappelant le granum des plantes supérieures et sont accolés sur une grande distance
C Le stroma et annexe plastidiaux.
Le stroma a un rôle métabolique et physique important car il renferme plusieurs enzymes, de l’ADN et des ribosomes. Les ribosomes des plastes sont différents de ceux des mitochondries. Parmi les annexes, on a les pyrénoïdes qui sont important quand on fait de la détermination d’algue. Ce sont des organites de nature protéique. Ils apparaissent comme différenciation du stroma plastidial. Chez les algues vertes, les pyrénoïdes sont entourés d’une membrane.
On a différentes localisations des chloroplastes : le stigma que l’on retrouve chez les cellules mobiles et il provient généralement de la différenciation d’une petite partie d’un plaste unique mais il peut également provenir de la transformation totale d’un plaste quand la cellule en possède plusieurs. Il est situé au voisinage de l’insertion des flagelles et est souvent associé à un photorécepteur pouvant être situé sur un renflement de la base flagellaire.
D L’appareil cinétique.
Beaucoup d’algues unicellulaires ou coloniales ont des flagelles. Le nombre de flagelles est souvent deux (on en a en général entre 0 et 4). Toutefois, on en trouve beaucoup plus chez certaines cellules reproductrices. Ces flagelles peuvent être égaux ou inégaux. Ils ont en général la même orientation. Les flagelles peuvent posséder des expansions fibrillaires appelées «mastigonèmes ».
Le nombre, la forme et la disposition des flagelles sont une des manifestations de la diversité des algues. Cela permet de caractériser des groupes ou des espèces dans un but taxonomique.
IV Les cycles de reproduction.
A Les rhodophytes.
Ce sont les formes les plus primitives ; elles proviendraient d’une seule symbiose. Elle a un chloroplaste à 2 membranes : la membrane d’une cyanobactérie plus la membrane d’une vacuole. Elles ont des points communs avec les cyanobactéries. On note la présence de billiprotéines (phycobilline) et de phycobillisomes situés à l’extérieur des thylacoïdes. Il n’y a qu’une seule classe : les rhodophycées qui sont pour la plupart des algues marines. Il y a 600 genres connus dont 29 en eau douce. Les algues marines sont presque toutes rouges alors que les formes d’eau douce le sont rarement (les billiprotéines sont solubles dans l’eau).
1 Caractéristiques.
- Elles possèdent de la chlorophylle A et D, donc possèdent des billiprotéines qui leur permettent de balayer tout le spectre d’absorption de la lumière, ce qui leur permet de pouvoir effectuer la photosynthèse. Elles vivent et se développent en grande profondeur. Leurs réserves sont constituées par l’amidon florigueen (ou rhodamylon).
- On peut observer la présence de synapses qui sont les stigmates laissés après une ouverture de la paroi lors de la formation de 2 cellules. Ces synapses ont aussi un rôle dans la migration de substances dissoutes qui est encore mal défini.
2 Morphologie.
Elles sont cladomientes, c’est à dire, typiquement constituées par un axe primaire non chlorophyllien. Cet axe primaire peut se ramifier en cladome II et III. Ces cladomes portent des rameaux courts, ramifiés à croissance limitée et sont toujours chlorophylliens : ce sont les pleuridies.
On observe différentes structures de cladomes :
- Des cladomes uniaxiaux à croissance indéfinie par le jeu d’une cellule initiale apicale. Ils portent des pleuridies alternes ou opposées. L’axe du cladome est toujours constitué par une seule file de cellules qui sont haplostichées.
- Des cladomes multiaxiaux avec des pleuridies typiques, non cortiquantes. L’axe cladomien est formé de plusieurs files de cellules.
Les cladomes uni ou multiaxiaux, avec leurs pleuridies, forment un cortex appliqué sur l’axe.
Toutes les cellules coxales portent des pleuridies, mais celles-ci sont très courtes, soudées, constituants des nœuds le long des filaments axiaux.
Les cladomes rhodoméloïdes sont toujours liés à une structure uniaxiale. Les pleuridies ont un grand développement et forment un cortex appliqué sur l’axe. Une seule n’est pas appliquée et forme la pleuridie chlorophyllienne. Elles sont distribuées comme les feuilles d’un phanérogame.
Quand le cladome est en lames folliacées, les pleuridies sont soudées entre elles.
3 Reproduction sexuée sans flagelle.
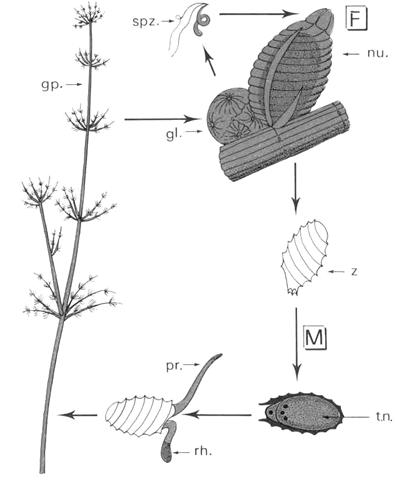
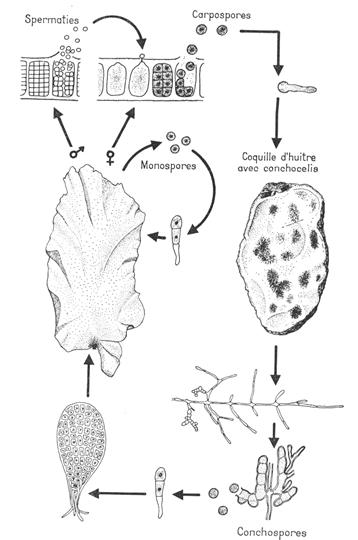
Les rhodophycées sont des algues marines de petite taille (1 à 4 cm), vivant sur les rochers et se développant sur les côtes de la Manche, l’Atlantique Nord et la Méditerranée. C’est une algue annuelle qui fructifie du printemps à l’automne.
Le gamétophyte est dioïque, bien que les plantes mâles et femelles soient haploïdes.
- Sur les thalles femelles, à partir de la cellule coxale d’une pleuridie, naît un rameau particulier : le rameau carpogonal. Il y a trois cellules incolores à la base du carpogone où de trouve l’oosphère munie d’un trichogyme.
- Sur les thalles mâles, à partir des cellules des pleuridies, naissent des petites ramifications portant de nombreux gamétocystes mâles produisant chacun un gamète unique nue et non flagellé : c’est une spermatie. Les spermaties flottent passivement dans l’eau et se fixent sur le trichogyme du carpogone et le contenu de la cellule de la spermatie passe du trichogyme puis au carpogone. A ce moment, il y a union des 2 noyaux. Le zygote ainsi formé est entouré de la paroi du carpogone surmonté du trichogyme qui flétrie. Les noyaux se divisent par mitoses successives. Un ensemble de cellules arrondies est un gonimoblaste. Ces cellules sont des carposporocystes qui donnent naissance à des carpospores avec un noyau à 2N.
L’ensemble carpogone plus gonimoblaste forme le carposporophyte. Ce dernier est situé sur le thalle qui lui a donné naissance. Ce qui donne un thalle à 2N, morphologiquement identique à ceux à N chromosomes. Ce thalle à 2N constitue le tétrasporophyte. Celui-ci donne naissance a des tétraspores provenant de cellules spécialisées : ce sont les tétrasporocystes où a lieu la méiose qui donnera 4 tétraspores à N chromosomes (espèce monoïque).
Les rhodophycées ont un cycle haplodiplophasique trigénétique (thalle à N chromosome donne un gamétophyte qui lui donnera un carposporophyte). Le gamétophyte donne le tétrasporophyte qui libérera les tétraspores.
B Les chromophytes
1 Généralités.
Ce sont des algues qui possèdent les chlorophylles A et C. Ils font parti de la classe des Phéophycées. Ce sont des algues en général marines. Leur taille et leur abondance leur donne un rôle important dans la végétation marine et dans les zones de balancement des marées. De plus, ces algues abritent une faune variée de poissons, crustacés qui y trouvent une nourriture abondante (c’est la chaîne alimentaire des bords de mer).
Par exemple : - au Japon, certains bords de mer sont exploités pour l’alimentation humaine : ce sont les Kombu. – Ils sont utilisés dans l’industrie alimentaire pour l’extraction des alginates dans : les yaourts, les cosmétiques, la peinture, l’imprimerie… Ils servent d’épaississants ou de gélifiants.
Dans ce groupe, l’anatomie et les modes de reproduction sont variés. Toutefois, les phéophycées possèdent une grande homogénéité dans leurs structures cytologiques. Leurs plastes contiennent de la chlorophylle A et C, et de la fucoxanthine (pigment spécial de coloration noire). Elles sont toujours pluricellulaires. Les cellules reproductrices mâles sont toujours biflagellées (un flagelle antérieur et un postérieur). Les flagelles s’insèrent sur le côté de la cellule.
2 Les cycles de reproduction.
On en a de 2 types : haplodiplophasique chez Ectocarpus et Laminaria, et diplophasique chez Fucus.
a Ectocarpus siliculosus.
Ils sont en forme de petites touffes de filaments bruns qui sont ramifiés et constitués de simples fils de cellules (ils sont rampants ou dressés). Chaque cellule renferme plusieurs plastes rubanés, où l’on trouve des pyrénoïdes.
On a deux types d’individus identiques morphologiquement :
- les gamétophytes à N chromosomes.
- les individus sporophytiques à 2N chromosomes.
1 La multiplication asexuée.
Elle se fait à partir de sporocystes pluriloculaires. Ils se forment à partir d’une cellule, qui subit de nombreuses mitoses, puis donne des petites loges qui vont donner naissance à une zoospore qui se fixera pour donner naissance à un nouveau gamétophyte. Si les zoospores sont haploïdes, ils proviennent d’un gamétophyte et s’ils sont diploïdes, ils proviennent d’un sporophyte.
2 La reproduction sexuée.
Certaines cellules issues des sporocystes, à partir des gamétophytes (mâles ou femelles) donnent des spermatozoïdes. Certaines gamètes se comportent comme des gamètes femelles attirants les gamètes mobiles mâles. C’est une reproduction de type planogamie car les gamètes mâles ont des flagelles (sont mobiles), isogame morphologiquement (même forme) mais anisogame fonctionnellement.
Les gamétophytes sont dioïques, qu’ils soient mâles ou femelles.
Le zygote formé va se développer en redonnent un sporophyte identique au gamétophyte. Dans des cas exceptionnels, le sporocyste reste sous la forme d’une grande cellule dont le noyau subit de nombreuses divisions cellulaires. Les nouveaux génomes donneront des gamètes.
Cycle haplodiplophasique, espèce dioïque, fécondation par planogamie isogame morphologiquement et anisogame fonctionnellement. La multiplication asexuée est réalisée grâce à des zoo-mitospores haploïdes et diploïdes.
b Les laminaires.
1 Généralités.
Ils peuvent mesurer jusqu’à quelques dizaines de mètres. Leur thalle est constitué par un stipe. La croissance se réalise entre le stipe et la fronde grâce aux méristèmes intercalaires. L’appareil végétatif diploïde donne le sporophyte. Il n’y a pas de multiplication asexuée par mitospores. Les trois parties du thalle (stipe, méristèmes et fronde) ont la même organisation histologique. De l’intérieur vers l’extérieur, on distingue :
- Le méristoderme. Il assure la croissance en épaisseur et produit uniquement des cellules vers l’intérieur, ce qui forme des couches concentriques rappelant les cernes du bois de printemps et d’automne (vers l’extérieur, on a les canaux mucifer). Seules les cellules externes sont chlorophylliennes. Tout le méristoderme intervient dans l’absorption des nutriments.
- Le cortex. Il est plus ou moins épais selon l’âge. Il est constitué d’assises de cellules séparées par une matrice intercellulaire
- La moelle ou zone médulaire. Elle est composée de files de cellules ramifiées à plastes peu nombreux. Ils forment des hyphes comme chez les mycètes. Les parois longitudinales sont plus épaisses chez les fibres (vaisseaux conducteurs).
2 La reproduction sexuée.
Quand l’appareil végétatif est fertile, il se couvre de grandes plages irrégulières appelées les «sores » qui sont plus sombres et légèrement en relief à la surface de la fronde. On trouve des sporocystes qui sont dressés perpendiculairement à la fronde et mélangés à des cellules stériles ou paraphyses.
Dans les sporocystes, s’effectue la méiose. Chaque sporocyste donne naissance de 32 à 64 méiospores biflagellés qui vont germés en donnant un nouvel organisme beaucoup plus petit : C’est le prothalle. Il est constitué de filaments rampants et de quelques filaments dressés et ramifiés. Sue ces derniers filaments se différencient les gamétocystes. On a deux types de gamétophytes (mâle et femelle). Les femelles sont plus grandes que les mâles. Les gamétophytes mâles portent les gamétocystes mâles qui produisent un gamète mâle chacun porteur de deux flagelles à insertion latérale. Les gamétophytes femelles portent les gamétocystes femelles à l’intérieur desquels il y a un gamète immobile : l’oosphère. Celle-ci n’est pas complètement libérée. L’oogame s’ouvre mais reste fixé au gametophyte.
Le zygote formé va redonner un appareil végétatif à 2N. Chez les laminaires, le cycle est digénétique (2 générations).
Le gamétophyte haploïde est de taille réduite et de durée de vie très courte. Il n’y a pas de multiplication asexuée chez les laminaires.
c Fucus vesicu.
C’est algue brune très répandue dans les mers tempérées et froides de l’hémisphère Nord.
1 Appareil végétatif.
Il est diploïde, constitué d’un ensemble de lanières plus ou moins rubanées, ramifiées dicotomiquement dans un même plan. L’algue est fixée sur un rocher grâce à un disque adhésif et peut atteindre quelques décimètres de long. Sur la fronde, on observe une nervure médiane saillante, des vésicules pleines de gaz (des flotteurs). La croissance de la fronde se fait par une cellule initiale unique, située dans une invagination au sommet des ramifications. Comme chez les Laminaires, il n’y a pas de multiplication asexuée.
2 La multiplication sexuée.
Le thalle est diploïde, c’est un sporophyte, qui porte des gamétophytes regroupés à l’intérieur de conceptacles qui sont à l’extrémité des frondes. Le fucus est une espèce dioique (une plante mâle et une plante femelle).
- Le gamétophyte mâle. A son extrémité, il y a des poils non fertiles (les paraphyses), qui sont de petits filaments ramifiés qui portent les gamétocystes. Chaque gamétocyste subit la méiose et donnent quatre noyaux. Puis, il subit quatre mitoses qui donnent 64 spermatozoïdes biflagellés sur le côté.
- Le gamétophyte femelle. On y trouve des réceptacles mélangés à des paraphyses non ramifiés, ce qui donne un gamétocyste femelle où se réalise la méiose qui va donner quatre cellules, puis huit oosphères non flagellées. Ces dernières sont libérées dans l’eau de mer et attirent les spermatozoïdes. Là, a lieu la fécondation qui donne naissance à un zygote à 2N qui germera en donnant un thalle mâle ou femelle à 2N. C’est une reproduction par oogamie. Le cycle est diplophasique et monogénétique, l’espèce est diplophasique.
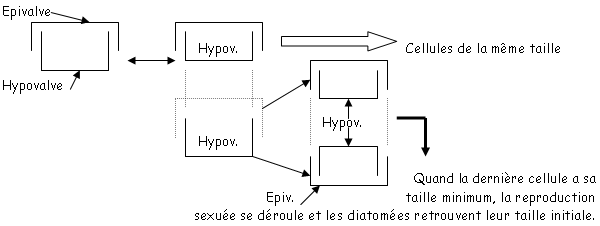
d Bacillarophyceae.
C’est une diatomée avec un important rôle écologique : ils constituent la base de chaînes écologiques et ont un rôle d’auto-épurateurs dans les rivières et servent d’indicateurs de pollution.
Ces diatomées sont constituées par 2 valves (épivalves et hypovalves). Quand la cellule se divise, il apparaît entre les deux valves, des connectives (ou ceintures ou bandes) qui sont appelées des cingulum. L’épivalve génère une valve de même dimension que son ancienne hypovalve. La diatomée est comme une boite de camembert.
C Les chlorophycées.
1 Généralités.
Ce sont des algues qui possèdent de la chlorophylle A et B.
Elles sont regroupées en quatre classes :
- Les chlorophyceae
- Les Prasinophyceae
- Les Zygophyceae
- Les charophyceae
On trouve, dans ces quatre classes, tout types d’algues : unicellulaires, flagellées, filamenteuses, ramifiées, en siphon (pas de cellules déterminées mais avec un grand nombre de noyaux)… Dans la sous-classe des chlorophycidées, on trouve par exemple l’ordre des volvocales, et comme type de chlorophycées flagellées, on a le genre Chlamydomonas.
2 Cycle de Chlamydomonas isogame.
C’est une espèce dioïque. Les cellules normales peuvent donner des sporocystes qui donneront des sporocystes qui redonneront des Chlamydomonas.
Le gamétocyste : il est obtenu grâce à une réunification des parties antérieures (là où sont les flagelles). Il donne un pré-zygote à quatre flagelles (pendant peu de temps). Ce planogamète perd ses flagelles et donnent un zygote avec une membrane épaisse, qui peut servir de forme de résistance. Dans le zygote, il y a la réduction chromatique qui donne les méiospores qui redonneront un nouveau Chlamydomonas. Cette espèce a un cycle monogénétique.
3 Cycle de reproduction de l’Ulve (Ulva lactuca).
L’Ulve fait parti de l’Ordre des Ulvales et de la sous-classe des Ulotrichophycidées. Le thalle est en forme de lame avec deux couches cellulaires. Il se reproduit à grande vitesse à cause de l’eutrophisation des côtes (les eaux marines sont trop riches en phosphore). Ces algues sont bien étudiées pour de multiples raisons :
- Elles n’ont pas de multiplication asexuée.
- Leur reproduction sexuée est celle d’une espèce dioïque.
Les deux types de gamétophytes sont identiques morphologiquement. Les cellules qui bordent ces thalles deviennent des gamétocystes et libèrent des gamètes biflagellés (de 16 à 32 gamètes pour le mâle et de 8 à 16 pour la femelle). Les gamètes femelles sont aussi biflagellées mais bien plus grosses.
La copulation donne un œuf planozygote à quatre flagelles. Il se fixe et donne un thalle diploïde, morphologiquement identique aux thalles haploïdes des deux sexes. Les cellules en bordure de ce thalle diploïde (ou sporophyte) donnent des sporocystes à l’intérieur desquels il y a réduction chromatique et formation de quatre méiospores quadriflagellées. Ceux-ci se fixent et redonnent des thalles haploïdes. Dans chaque sporocyste, on a deux méiospores mâles et deux femelles.
Cycle haplodiplophasique, digénétique (à deux générations). Comme il a des thalles mâles et femelles, individus dioïques et planogamie anisogame car les gamètes mâles et femelles se différencient par leur taille.
4 Cycle de Oedogonium.
Il fait toujours parti des chlorophycées mais de la sous-classe des Oedoniophycidées.
Cette sous-classe se caractérise par des genres coloniaux qui forment des filaments. Ces filaments se caractérisent par une croissance intercalaire : seules certaines cellules se divisent. Celles qui ont subi la division portent une calotte marquée par des cicatrices (on a la formation d’un bourrelet). Les espèces de Gedogonium sont monoïques ou dioïques.
Pour reconnaître une espèce, il faut observer le zygote. Les zygotes portent différentes ornementations.
Dans le cas des monoïques, le sporocyste va former une zoospore (ou androspore) qui va s’insérer dans la cellule se trouvant sous l’oosphère puis va former un mâle nain qui, lui, ira féconder l’oosphère.
5 Cycle des zygophycées. Cas du Zygnéma.
Ce sont des algues filamenteuses ou unicellulaires. Leur caractère commun est la façon dont se déroule la reproduction sexuée.
Le zygnéma est une espèce dioïque. La reproduction a lieu quand les conditions du milieu deviennent défavorables. Les filaments se mettent en parallèle les uns aux autres. Une cellule prend le rôle de cellule mâle et fait migrer tout son cytoplasme vers la cellule contiguë (réceptrice ou femelle). Il y a fécondation et formation d’un zygote par une cystogamie anisogame. Quand une cellule commence à subir ce phénomène, toutes les autres font de même : c’est une conjugaison scalariforme. Il n’y a pas de gamètes flagellés. L’œuf s’entoure d’une membrane épaisse, différemment ornée selon les espèces, puis se laisse tomber au fond du milieu de vie. Quand les conditions redeviennent favorables, il germe, subit la méiose et donne quatre noyaux haploïdes dont trois qui dégénèrent. Le noyau subsistant va donner un nouveau thalle haploïde.
Remarque : Pour la cystogamie isogame, le contenu des deux cellules migre dans un canal copulateur où le zygote se forme.
6 Les pyrophycées.
Elles ont deux parties séparées par sillon longitudinal où se trouve un flagelle. Un autre flagelle se trouve dans un autre sillon partant du centre.
Chapitre 4 : Les Champignons.
I Généralités.
C’est une espèce immobile qui a longtemps été classée chez les végétaux mais qui a des similitudes avec les animaux (mode de vie, paroi cellulaire en chitine).
Maintenant, les champignons ont un règne propre : les Fungi. C’est un vaste ensemble d’organismes eucaryotes (300.000 espèces connues) mais on estime la totalité à 1,5 millions d’espèces.
Ils sont souvent sous forme filamenteuse pluricellulaire. On y trouve tous les modes de nutrition (saprophytes, parasites, symbiontes mutualistes). Il y a beaucoup de variations de structures et de fécondations. Ce sont des organismes hétérotrophes pour le carbone : ils dépendent de molécules préformées, pour croître et se reproduire. Ils sont dépourvus de chlorophylle et ne peuvent donc pas réaliser la photosynthèse. Ils sont capables de dégrader des molécules complexes pour obtenir l’énergie et le carbone nécessaire. Ils ont une importance économique dans : l’agro-alimentaire (alcool, pain, fromage, carpophores), la pharmacologie (les antibiotiques. Ils sont responsables de maladies chez les plantes (mildiou…). Ils peuvent détruire des charpentes en bois (mérules…), sont parfois toxiques et peuvent entraîner des mycoses.
Ils sont un maillon essentiel des écosystèmes (décomposition de la matière organique, recyclage des éléments, constitution de réseaux trophiques). Ils favorisent ou limitent la dispersion de leur hôte quand ils sont parasites. On les trouve en milieux aquatiques, mais surtout terrestres.
II L’appareil végétatif : Cycle de vie.
En germant, les spores forment des hyphes (tubes de petit diamètre, entre 5 et 10 µm) qui se ramifient et donnent un réseau complexe de filaments : le mycélium. C’est un organisme modulaire dont l’unité de base est l’hyphe. La construction du mycélium se fait par répétition de cet hyphe. Les filaments sont cloisonnés ou non. Quand les filaments sont cloisonnés, ils donnent un mycélium siphonné (coenocytique). Il y a formation d’une cloison entre le gamétophyte (ou le sporocyste) et le reste du thalle. Quand les filaments ne sont pas cloisonnés, il y a des parois entre cellules (ou articles (avec plusieurs noyaux)). Ce sont ces formations qui donnent la rigidité du thalle mais le flux cytoplasmique est toujours possible.
Parfois, on a des regroupements de filaments qui forment des amas (c’est le blanc des champignons) ou des tissus blancs (les plectenchymes). Les thalles sont parfois très ramifiés. Parfois, les champignons alternent leurs formes : filamenteuse puis unicellulaire et inversement. Parfois, les thalles constituent un plasmode (masse de cytoplasme multinucléé), sans paroi squelettique (exemple : la hernie du chou avec Plasmodiophora brassicae).
III Variétés des modalités de reproduction.
Les champignons sont capables d’assurer leur dispersion vers de nouveaux habitats et de s’y installer tout en étant plus ou moins en contact avec le substrat d’origine, car ceux-ci sont non-mobiles. Ils se dispersent vers de nouveaux habitats grâce à une extrême diversité de modalités de reproduction et un grand pouvoir de dissémination et de multiplication.
Les spores sont de très petite taille. Leur transport est passif et a lieu sur de grandes distances grâce au vent et à l’eau. Leur dispersion est aussi possible grâce aux insectes par la production de spores enveloppées de sécrétions sucrées ou odoriférantes (exemple avec Phallus impudicus).
A La reproduction végétative (multiplication asexuée).
C’est la production de nouveaux individus, identiques à l’organisme qui les a produit. La genèse des spores se fait selon différents mécanismes :
- Fragmentation de l’hyphe : formation d’arthrospores.
- Formations de bourgeonnements en chaînes : ce sont les blastospores.
- A l’intérieur de sporocystes
- Par des cellules spécialisées : les philiades.
La formation de spores exogènes (conodies), est caractéristique des ascomycètes, basidiomycètes et deutéromycètes.
B La reproduction sexuée.
Elle implique la fusion de cellules haploïdes produites par des organismes d’une même espèce mais de caractères génétiques différents. L’œuf (ou zygote) possède un double jeu de chromosomes : il est diploïde.
Il y a un double brassage génétique : les cellules qui fusionnent ont une composition génétique différente. Au cours de la réduction chromatique, il peut y avoir échange d’ADN entre les chromosomes homologues.
Ces nouvelles entités sont différentes des types parentaux et sont donc génétiquement uniques.
La fécondation se déroule selon diverses modalités, elles sont différentes d’un groupe à l’autre et entre deux espèces. S’il y a fusion entre gamètes mâles et femelles bien différenciés : c’est la gamétogamie. S’il y a fusion de gamétocystes (sans gamètes) : c’est une cystogamie. Chez certains champignons, il y a fécondation entre deux cellules sans différenciation particulières : c’est la somatogamie.
IV Embranchement des champignons : classification.
La classification est basée sur la composition des parois, sur la structure des filaments et des organes reproducteurs. Les groupes sont hétérogènes, les ancêtres sont différents mails pas le mode de vie.
A Les Myxomycètes (500.000 espèces).
Ils sont beaucoup plus proches des protozoaires que des autres champignons. Leur mode de vie et leur organisation sont plus plutôt protozoaires et leurs modalités de reproduction sont proches de celles des autres champignons.
1 L’appareil végétatif.
C’est un plasmode : une masse de cytoplasme unique et visqueux pourvu de nombreux noyaux et sans paroi squelettique. Leur mobilité est de type amiboïde. Ils font quelques centimètres d’épaisseur.
Leur organisation est coenocytique.
2 L’appareil reproducteur.
Sa mise en place s’accompagne de la mise en place de cloisons.
B Les oomycètes.
Ce sont des cellules mobiles avec deux flagelles, à paroi cellulosique. Les filaments siphonnés sont coenocytiques. Leur habitat est aquatique, mais un groupe, les péronosporales, s’est adapté à la vie terrestre en devenant parasites de plantes.
1 Cycle de Plasmopara viticola.
C’est un parasite de la vigne qui se trouve sur les feuilles et les grappes. Dans les tissus de la vigne, ils développent un mycélium diploïde qui envoie des suçoirs dans les cellules hôtes. Ce parasitage rend les feuilles jaunâtres.
a La multiplication asexuée.
Quand le temps est humide, le mycélium de la face inférieure de la feuille fait sortir des filaments par les stomates. A l’extérieur, les ramifications sont perpendiculaires aux filaments : ce sont les stérigmates. A l’extérieur, ces derniers se développent en sporocystes. Il y a séparation des filaments par un cloisonnement, ce qui fait tomber les sporocystes et leur permet d’être disséminés. Ils se déposent sur les organes verts de la vigne. S’ils tombent dans l’eau, ils donnent des zoospores qui vont donner des filaments à 2N qui se répandent grâce à plusieurs multiplications en une saison.
b La reproduction sexuée.
A la fin du cycle de végétation, dans les tissus hôtes, apparaissent les organes sexués (gamétocystes) où s’effectue la méiose. Dans les gamétocystes mâles, il n’y a pas d’individualisation. Dans les gamétocystes femelles, l’oocyste se scinde en deux zones : le périplasme (la zone externe) et l’ooplasme (la zone interne).
Le gamétocyste mâle va s’accoler au gamétocyste femelle : formation d’un tube copulateur qui est utilisé par un noyau mâle pour aller dans l’oocyste. Il y a cystogamie avec siphonogamie.
Quand l’ooplasme est fécondé, il y a apparition du zygote avec une paroi épaisse qui assure la conservation de l’espèce pendant l’hiver. Au printemps, les zygotes germent en émettant des filaments, au bout desquels on trouve les sporocystes qui libèrent leurs spores biflagellées à 2N. Ce sont ces spores qui sont à l’origine de la nouvelle contamination.
Le cycle est diplophasique. L’espèce est monoïque. La fécondation est une cystogamie avec siphonogamie. La multiplication asexuée se fait par des spores à 2N.
2 Le polyplanétisme avec Achlya ambisexualis.
Cette espèce est dioïque avec des thalles diploïdes. La multiplication asexuée se réalise avec des filaments dont l’extrémité se renfle pour donner les sporocystes qui vont se séparer du reste du mycélium par une cloison. Ils vont donner des zoospores biflagellées qui s’enkystent immédiatement. Ces spores vont redonner des zoospores flagellées (flagelles latéraux antérieures). Plusieurs enkystement sont possibles : c’est le polyplanétisme.
C Les chytridiomycètes.
Ces champignons représentent à peu près 1000 espèces. Leur caractéristique distinctive est que leurs cellules sont monoflagellées pendant une courte durée du cycle.
Le thalle : il est non-cloisonné (coenocytique). Souvent, la caryogamie succède à la plasmogamie. Ils sont considérés comme de vrais mycètes car leurs parois contiennent de la chitine et des sucres de réserves (le glycogène). C’est le groupe qui a du donner naissance aux groupes plus évolués. L’habitat est souvent aquatique.
Les zoospores se fixent souvent sur des diatomées par leur «ceinture » et se transforment en une cellule qui donnera un sporocyste.
Par moments, les zoospores donnent des gamètes mâles et femelles qui se fixent sur les diatomées en donnant des gamétocystes mâles et femelles. Les gamètes mâles passent dans les gamètes femelles par cystogamie avec siphonogamie. Le zygote formé est à 2N et après avoir subit la méiose, il donne des zoospores à N qui se fixent à leur tour sur la diatomée
D Les zygomycètes, avec le cas de Mucor mucedo.
Dans cette espèce, il y a absence de cellules mobiles. Le thalle est siphonné et coenocytique, et présente une zygospore à 2N à paroi très résistante (conservation). Cette zygospore résulte de la fusion de deux gamétocystes compatibles sexuellement : c’est une fécondation par cystogamie.
La multiplication végétative est réalisée par les spores produites à l’intérieur de sporocystes pédicellés. Parfois, la multiplication végétative se réalise par fragmentation du thalle.
L’habitat de Mucor mucedo est terrestre. C’est la moisissure du pain.
1 La multiplication asexuée.
Sur le mycélium, se dressent des filaments au bout desquels se trouve une petite boule noire : ces boules sont les sporocystes qui sont supportés par un sporocystophore qui se prolonge dans le sporocyste par une columelle. Le contenu du sporocyste se fragmente en un grand nombre de spores (sans flagelle) qui germent en redonnant un mycélium haploïde.
2 La reproduction sexuée.
Les gamétocystes se forment à partir d’un mycélium haploïde. Ils s’individualisent à l’extrémité de ramifications qui se renflent. Ils se séparent du filament porteur (ou suspenseur) par un cloisonnement. Les gamétocystes sont attirés l’un par l’autre et fusionnent en donnant un zygote. Il faut toutefois que ces gamétocystes soient compatibles (hétérothallisme). L’œuf s’entoure d’une membrane épaisse et est formé de plusieurs noyaux. Dans un premier temps, les noyaux se multiplient puis fusionnent (+ par -). Ceux qui ne fusionnent pas dégénèrent. Ici, tous les diploïdes dégénèrent sauf un qui subit la méiose et donne quatre cellules haploïdes (deux + et deux -) dont seulement deux survivent et redonneront un nouveau thalle.
Le cycle est haplophasique, hétérothallique. La fécondation est une cystogamie et la multiplication asexuée se fait par spores non flagellées.
E Les ascomycètes.
C’est le groupe le plus important des champignons, si on compte les lichens et les formes classées chez les deutéromycètes : 30.000 espèces. C’est aussi le groupe le plus diversifié et il présente de grandes variétés d’appareils reproducteurs, de modes de vie et de dissémination. Ils sont d’une grande importance économique. Ce sont : les parasites de cultures (pourritures grises des fruits et légumes : l’oidium) ; la maladie hollandaise de l’orme ; de bons mycètes.
Il y a la présence d’un asque : c’est un méiosporocyste dans lequel à lieu la méiose. Le mycélium filamenteux est haploïde, très ramifié et cloisonné. Il y a complexification des formes morphologiques avec souvent, la formation de faux tissus ou de plectenchymes.
La fécondation est une caryogamie différée. Ce sont de vrais champignons. Leur multiplication végétative est très répandue.
Cas de Neurospora crassa.
C’est un saprophyte d’excréments. Il est utilisé comme modèle dans de nombreuses recherches scientifiques.
1 La multiplication asexuée.
L’extrémité de certains filaments donne des spores ellipsoïdales qui contiennent plusieurs noyaux. Ces exospores (ou conidies) sont dispersées par les insectes, le vent et ils germent en donnant un nouveau thalle haploïde.
2 La reproduction sexuée.
La fécondation présente une convergence morphologique comme chez les algues rouges. Sur le mycélium, se dresse un filament particulier (le filament ascogonial) qui est constitué de grosses cellules plurinucléées (ou articles). A la base de ce filament, se développe un autre filament qui donne le proascocarpe. Le filament ascogonial est surmonté du trichogyme. L’organe femelle est l’association de l’ascogone et du trichogyme (le trichogyme joue le rôle de capteur de gamètes mâles). Les gamètes mâles ne naissent pas de sporocystes, mais elles sont bourgeonnées latéralement par les articles de certains filaments. Les spermaties n’ont pas de flagelles et ont un noyau volumineux. Elles sont emmenées passivement vers le trichogyme. Quand il y a contact, le noyau migre vers le trichogyme jusqu’à l’ascogone qu’il féconde. Plusieurs spermaties peuvent fusionner avec le trichogyme mais une seule va féconder l’ascogone.
3 Le développement du zygote.
L’ascogone fécondée est diploïde Le développement se fait sur place et engendre des filaments présentant la particularité d’être divisés en cellules comptant deux noyaux chacune. Ces sont des cellules à dicaryons.
On observe le phénomène du crochet : l’extrémité d’un dicaryon bourgeonne latéralement. Les deux noyaux subissent une division et un noyau fils passe dans le bourgeon alors que l’autre reste dans la première cellule. Le bourgeon latéral se sépare de la cellule terminale par une cloison. On obtient trois cellules : une cellule à deux noyaux différents et deux cellules à un noyau. : c’est une dangeardie. Le bourgeon se recolle à la cellule initiale et y fait passer son noyau (deux cellules à deux noyaux distincts).
4 La formation de l’asque.
La cellule de l’asque est le siège d’une méiose. Les filaments dicaryontiques se terminent par une cellule où les deux noyaux fusionnent en un noyau diploïde. C’est la cellule ascogène qui va s’allonger alors que le noyau (à 2N) subit la méiose. A la fin, on a huit noyaux haploïdes (quatre + et quatre-) autour desquels s’individualise une ascospore. Un asque est l’association d’une membrane et de huit noyaux haploïdes. Ces ascospores vont donner un nouveau mycélium haploïde.
5 La formation de l’ascocarpe.
Les filaments qui entourent l’ascogone (le proascocarpe) se développent et constituent la paroi d’un conceptacle ouvert à la partie supérieure par un ostiole. C’est la formation de l’ascocarpe. Un ascocarpe a une morphologie variable : globuleux, ouvert par un pore ou non (c’est alors un périthèce) ou en forme de coupe (apothécie). Les ascocarpes peuvent être constitués par une masse de tissus avec des veines. Dans le cas des levures, on n'a pas d’ascocarpe.
Ici, le cycle est haplodiplophasique. La fécondation est une trichogamie. La multiplication se fait par les conidies.
F Les basidiomycètes.
Ce sont les plus connus. Ils forment de gros carpophores très visibles. Ils ont des conséquences économiques : ils sont parasites de cultures et d’arbres (rouilles, charbons), ils peuvent aussi être saprophytes (destructeurs de charpentes).
Ces champignons possèdent des basidies (l’équivalent d’un méiosporocyste) où se réalise la méiose. Le mycélium filamenteux est très ramifié et cloisonné. On a souvent des formations de faux tissus (pectenchymes). Il y a complexification des formes morphologiques. La fécondation se réalise par une caryogamie différée. Le mycélium secondaire (N+N) constitue la phase dominante du cycle. La fécondation est une somatogamie (thallogamie). Ce sont aussi de vrais mycètes. Il existe deux groupes : les hétéro et les homobasidiomycètes.
Les homobasidiomycètes peuvent être : saprophytes (champignons de Paris) ; symbiotiques (cèpe, amanite de César) ; parasites (les polypores comme la Langue de bœuf).
1 Cycle de Coprin chevelu ou Coprinus comatus, un homobasidiomycète.
On le trouve dans les jardins, les cultures, les bords de chemin (dans les endroits engraissés). On le trouve à la fin de l’été et pendant l’automne.
Le carpophore est la partie visible qui montre qu’il y a dessous, un mycélium.
a Développement et cycle.
Le stade haploïde se trouve dans le gamétophyte, à partir de basidiospores qui vont donner des filaments mycéliens à cellules uninucléées. C’est le filament primaire (+ ou -). Ils peuvent donner naissance à des spores extérieures qui peuvent disperser l’espèce au stade haploïde.
b La fécondation (somatogamie)
Entre deux filaments I + et I-, se produit une anastomose qui donne un mycélium II à dicaryon (noyaux + et -). Le noyau se divise par le phénomène du crochet.
Les filaments secondaires s’agrègent pour constituer un cordon sur lequel se différencient des boules blanches (ébauches de fructification ou carpophore). Sur les carpophores sont portées les basidies. A l’extérieur, on a la membrane blanche (le voile universel), à l’intérieur duquel se constituent les différentes parties. Dans ce voile, on a, le jeune pied, le chapeau dont les bords sont repliés et viennent s’unir au pied par le voile partiel. Pendant le développement du carpophore, le pied s’allonge et le voile universel se casse et ne persiste sur le chapeau que sous forme d’écailles. Le diamètre du chapeau augmente et le voile partiel se rompt en formant un anneau autour du pied.
Au niveau des lamelles, se différencient les basidies. En coupe, les lamelles sont constituées au centre par des éléments parallèles entre eux et recouverts de filaments emmêlés très denses (le sous-hyménium) recouverts par l’hyménium. On a des basidies perpendiculaires à la surface des stériles.
c La formation des basidies.
Les basidies se différencient à partir de cellules terminales du filament du sous-hyménium. Dans cette cellule pro-basidie à lieu la caryogamie (union des deux noyaux). La cellule croit et donne la baside où à lieu la méiose, ce qui entraîne la formation de quatre basidiospores (2+ et2-) qui sont bourgeonnées à l’extérieur de la basidie (par de petits pédicelles, les stérigmates) et germent en donnant un mycélium primaire (+ ou -).
Le cycle est haplophasique, digénétique. Il y a hétérothallisme. La fécondation est une somatogamie. La multiplication asexuée se fait par les conidies.
2 Les hétérobasidiomycètes, cas de Puccina graminis.
Ce sont des parasites des végétaux supérieurs dont la basidie est cloisonnée. Exemple : la rouille du blé : Puccina graminis. Puccina a deux hôtes différents : l’épine vinette (Berberis vulgaris) et le blé.
a Le stade haploïde et la Berberis.
Le stade haploïde est le gamétophyte. Le développement de ce mycète sous forme haploïde se réalise quand il est sur une feuille de Berberis. Il germe et donne un filament (à cellule à un noyau) et envoie des suçoirs dans les cellules foliaires de l’hôte. Ce parasitage entraîne une production de taches jaunes /oranges sur les feuilles. Les taches sur la face supérieure sont des pycnides ou spermogonies et sur la face inférieure se sont des écidies.
Les pycnides : sont organisés en forme de bouteille logées dans des cavités de la feuille ouverte vers l’extérieur. On observe deux sortes de filaments : les premiers se fragmentent en chapelets de cellules uninucléées (pycnidiospores). Leur dissémination est assurée par les insectes, ils ne germent pas (ils sont comme des gamètes mâles). Par l’ouverture du pycnide sort un filament sexuel en relation avec le mycélium haploïde : ce sont les hyphes récepteurs. La fécondation se fait par ces hyphes (ils ont le rôle de trichogyme).
A la face inférieure de Berberis, on a des enchevêtrements d’hyphes qui constituent des nodules (proécidies non fécondées), limitées par la paroi mycélienne (tissu nourricier et tissu fécondable).
Quand les spermogonies sont mûres, elles excrètent une goutte de nectar où se trouvent les spermaties transportées par les insectes vers d’autres feuilles. L’espèce est hétérothallique. Si la spermatie rentre en contact avec l’hyphe récepteur d’un mycélium de signe opposé, le noyau pénètre l’hyphe et passe d’une cellule à l’autre jusqu’au tissu fertile de l’écidie dont les cellules deviennent dicaryontiques. Dans un premier temps, les noyaux + et – ne fusionnent pas. Ce sporophyte se développe au dépend du tissu nourricier. Le développement donne naissance à un organe en forme de cupule : l’écidie mature. Sa taille augmente et casse l’épiderme de la face inférieure de la feuille. Un filament donne naissance à l’écidiospore à 2 noyaux, paroi épaisse avec une cellule disjonctrice. Les écidiospores ne germent que si elles rencontrent du blé. Là, il y aura formation d’un mycélium dicaryontique, entre les cellules du blé.
b Le stade diploïde et le blé.
Dans la feuille de blé, le mycélium donne des fructifications allongées sous-épidermiques : les urédosores. Sur les urédosores se différencient les urédospores binucléées, qui se trouvent à l’extrémité d’un long pédicelle. Elles font éclater l’épiderme, donnant à la feuille une couleur rouille. Les urédospores peuvent germer sur le blé, ce qui propage la maladie, avec formations de filaments à dicaryon
La multiplication végétative. Elle se réalise grâce à des téleutospores. Le mycélium dicaryontique constitue un autre type de fructifications de couleur noire (c’est un groupement sous forme de téleutosores, ensemble de pédicelles à l’extrémité desquels se trouvent les téleutospores). Ils sont dicaryontiques, tombent sur le sol où elles passent l’hiver. Pendant cette période, les noyaux fusionnent.
Au printemps, chaque cellule de la téleutospore émet un filament, le promycélium, où s’engage le noyau diploïde qui subie la méiose et donnent deux noyaux + et deux -. Ces quatre noyaux s’isolent par une cloison et donnent des basidies cloisonnées (les hétérobasidies). Chaque cellule émet un prolongement latéral (les stérigmates) qui bourgeonne une basidiospore où passe un noyau, puis celle-ci est libérée.
Cette basidiospore infectera ensuite la Berberis.
Le cycle est trigénétique, le gamétophyte est hétérothallique, saprophyte à dicaryon avec deux éléments. Le mycélium forme les écidies. Le mycélium à urédospores et téleutospores est indépendant du gamétophyte. La fécondation est par hyphe réceptrice. La multiplication végétative se fait par urédospores.
Les plantes supérieures : Généralités.
Les plantes supérieures sont des archégoniates, cormophytes, eucaryotes, faisant parti des végétaux. Elles possèdent un cormus constitué de rameaux feuillés (sauf les bryophytes et certaines ptéridophytes). Les archégoniates possèdent des racines et, pour la plupart, des tissus bien différenciés mis en place par les méristèmes primaires ou secondaires (ce sont des formations de cellules embryonnaires indifférenciées), à paroi fine, pouvant se multiplier activement.
Les gamètes sont renfermés dans les gamétanges (qui sont les anthéridies mâles ou les archégones femelles), qui sont toujours entourés par une enveloppe constituée d’au moins une assise de cellules.
On a plusieurs types de fécondation :
- Zoïdogamie : le gamète mâle est mobile et le gamète femelle est immobile (oogamie).
- Siphonogamie : le gamète femelle est immobile, le gamète mâle n’est ni flagellé ni cilié mais la fécondation se fait par un siphon ou par un tube pollinique.
Les spores sont formées dans les sporanges, toujours après une méiose.
Classification :
- Bryophytes (mousses)
- Ptéridophytes (fougères)
- Pré spermaphytes
- Spermaphytes :
- gymnospermes
- angiospermes : mono et dicotylédones.
Chapitre 5 : Les Bryophytes.
Certains bryophytes sont encore thalloïdes (avec des thalles) : ce sont les hépatiques. D’autres possèdent une tige et des feuilles mais pas de racines, seulement des rhizoïdes ayant un rôle de fixation et d’absorption.
Les bryophytes sont séparées des autres archégoniates par le fait qu’elles sont dépourvues de vaisseaux pour la conduction de la sève ; cependant, chez les espèces les plus évoluées, il y a un début de différenciation des tissus. Il existe une grande domination du gamétophyte par rapport au sporophyte qui est, toute sa vie, parasite du gamétophyte.
Les bryophytes vivent le plus souvent en milieu humide. On observe sur ces mousses le phénomène de reviviscence : elles peuvent supporter une dessiccation prolongée, en passant à un état de vie ralentie. Quand des conditions viables reviennent, elles font repartir leur métabolisme.
I La classe des hépatiques, avec Riccia sp.
A Le gamétophyte.
Au départ, on a un mitospores à N, puis il germe sur un sol humide et donne de petits filaments : des protonéma de 4 cellules chacun (une cellule est un rhizoïde). Le développement est réalisé par les divisions successives de la cellule apicale, ce qui donne un thalle prostré sur le sol. Ce dernier a l’aspect d’une rosette à plusieurs branches.
Sur le thalle, on peut distinguer deux zones :
- Le parenchyme ventral, riche en réserves et portant les rhizoïdes et les écailles (les amphigastes).
- La zone supérieure est formée par un parenchyme chlorophyllien ou assimilateur. On trouve de nombreux canaux aérifères. L’assise supérieure (ou épiderme) est non chlorophyllienne.
B L’appareil reproducteur de Riccia sp.
Il est composé des anthéridies et des archégones sur la partie dorsale du thalle, dans le sillon longitudinal.
1 Les anthéridies.
Elles sont formées par une cellule superficielle du thalle qui, par divisions successives, donne un sac formé par une enveloppe externe constituée d’une assise de cellules. A l’intérieur, les nombreuses cellules sont obtenues par mitose : c’est la différenciation en anthérozoïdes.
Il y a libération des méiospores grâce à la résorption de la paroi externe. Ces spores sont formées de gros noyaux et possèdent deux flagelles.
2 Les archégones.
C’est une cellule superficielle, qui, à maturité, a la forme d’une «bouteille » attachée au thalle par un pied pluricellulaire (ou pédicelle). Quand le développement de l’archégone est complet, le ventre est surmonté d’un col (ou calyptra) constitué d’une seule couche de cellules. L’oosphère est à l’intérieur du ventre.
3 La fécondation.
A maturité, l’archégone s’ouvre au sommet. Les cellules qui se trouvent à l’intérieur du col se gélifient. Les anthérozoïdes, attirés par chimiotachtisme (à cause des sucres et des substances de mucilage), se déplaçant dans une goutte d’eau, vont pénétrer dans le col et un seul des anthérozoïdes va s’unir à l’oosphère, ce qui va donner un œuf. C’est la première cellule (à 2N) du sporophyte dont la fécondation s’effectue par zoïdogamie
4 Le sporophyte à 2N.
L’œuf, par divisions successives, donne un sporophyte globuleux inclus dans le ventre de l’archégone. Il présente deux zones distinctes :
· une zone externe : l’amphithécium qui forme la paroi du sporophyte
· une zone interne : l’endothécium qui par mitoses va donner des cellules nourricières et les cellules mères des archéspores.
· les cellules mères subissent la réduction chromatique et chacune d’elles donnera 4 méiospores libérées après la destruction des tissus du thalle (en général, un an après).
· La nouvelle génération donne un protonéma.
Remarques :
· par liquéfaction, les cellules nourricières et l’amphithécium forment un liquide nutritif.
· les archéspores sont toujours dans le ventre des archégones.
Le sporophyte a une existence transitoire, c’est un parasite du gamétophyte. Il disparaît après la réduction chromatique.
Le cycle est haplodiplophasique mais avec une dominance du gamétophyte.
II La classe des mousses, avec Funaria hygrometrica.
On la trouve dans les sous-bois clairs, dans les landes après les incendies
A Le gamétophyte : l’appareil végétatif.
Une méiospore à N qui germe sur un sol humide et donne naissance à un filament chlorophyllien unisérié, rampant : le protonéma. Ce filament se ramifie et émet des rhizoïdes. Sur ces filaments naissent des cellules renflées qui après s’être divisées, donnent les tiges feuillées de la Funaria : c’est le gamétophore. Le protonéma disparaît et laisse un ensemble de gamétophores groupés. Chaque pied présente la seconde partie de l’appareil végétatif. Celle-ci est représentée par une tige parfois ramifiée au sommet, portant des petites feuilles à la base et des rhizoïdes unisériés. La tige possède une anatomie simple : un épiderme, une couche d’une assise ou deux de cellules de soutient (les stéréides), une zone corticale parenchymateuse. La zone interne est composée de cellules de fine section jouant un rôle dans la conduction de l’eau. Les feuilles de petites tailles (les microphylles) sont formées d’un limbe d’une seule assise de cellules. La nervure centrale présente des cellules superficielles épidermiques et des cellules internes qui sont des conducteurs.
B L’appareil reproducteur monoïque.
1 Les anthéridies.
Elles sont situées à l’extrémité des rameaux latéraux des gamétophores. Elles se trouvent dans des corbeilles spécifiques, dites «à anthéridies », fermées par un ensemble de feuilles modifiées par rapport aux autres : ce sont les feuilles périgoniales. Entre les anthéridies, se développent des poils stériles : les paraphyses. Les anthéridies se développent à partir d’une cellule superficielle. Des sacs allongés sont reliés à la corbeille par un pied bisérié, à l’extrémité se trouve l’opercule. Les cellules internes se divisent par méiose et donnent des anthérozoïdes qui sont à N chromosomes.
2 L’archégone.
Il y en a de 2 à 5 par pied. Elles sont situées à l’extrémité de la tige principale, entourées de feuilles périgoniales mais pas de paraphyses. Le développement à lieu à partir d’une cellule unique superficielle. Sa morphologie rappelle celle du Riccia sp. , mais le pied est plus épais et le ventre est constitué de 2 ou 3 assises de cellules.
La fécondation : A maturité, les cellules de canal du col (à l’extrémité supérieure) se gélifient. Les anthérozoïdes sont attirés par chimiotachtisme et se déplacent vers l’archégone en nageant dans l’eau de pluie ou dans de la rosée, puis ils pénètrent dans le col. Un anthérozoïde fusionne avec l’oosphère et donne un zygote entouré par une membrane cellulosique.
La reproduction est une zoïdogamie tributaire du milieu (eau ou rosée nécessaire).
3 Le sporophyte.
L’œuf grossit par mitose et donne le sporophyte (ou sporogone). Il se développe ensuite en entraînant l’archégone qui donnera naissance à la coiffe. Le sporophyte est constitué par le pied (suçoir) fixé en parasite sur le gamétophyte, la soie (long filament), la capsule (ou urne) qui est obturée par les dents du péristome et fermée par l’opercule et le tout est recouvert par la coiffe. Cette urne contient les méiospores à maturité.
4 L’urne.
Elle se différencie à partir d’une cellule provenant des divisions de l’œuf. Ces cellules (de l'urne) s’organisent en endothécium et amphithécium (constitué par un épiderme pouvant posséder des tomates, par un parenchyme lacuneux et chlorophyllien, par 2 ou 3 assises de cellules qui forment la paroi).
5 Les archéspores.
En se divisant, les archéspores donnent naissance à des cellules nourricières et aux cellules mères. Ces dernières subissent la réduction chromatique et donnent 4 méiospores (ou tétraspores). Lorsque ces spores sont mûres, la coiffe puis l’opercule tombent, les dents s’écartent, la soie se courbe et les spores peuvent être disséminées et donneront un protonéma quand le temps deviendra humide.
Comme chez Riccia, le gamétophyte est prédominant (le sporocyste est parasite), la fécondation est une zoïdogamie, le cycle est haplodiplophasique digénétique et l’espèce est diploïde.
Remarque : Les gamétanges qui donnent naissance à des assises cellulaires sont différents des gamétocystes des algues et des champignons qui donnent des «graines » séparées par la paroi.
Chapitre 6 : Les Ptéridophytes.
I Généralités.
Elles font parti de l’embranchement des archégoniates. Ce sont des cormophytes. Les gamètes sont formés dans les gamétanges. Les ptéridophytes sont des cryptogames vasculaires car ils possèdent des vaisseaux du bois de type scalariforme. Morphologiquement, on a une tige, des feuilles et des racines. Le sporophyte est la forme prédominante, chlorophyllien, rapidement autonome par rapport au gamétophyte. Ce dernier est souvent thalloïde et dépourvu de vaisseaux conducteurs.
Les ptéridophytes vivent en général dans les milieux humides, et certains sont aquatiques (ex : Azolla ). Certaines formes comme Ceterach peuvent résister à la sécheresse. Dans les forêts tropicales, les fougères sont arborescentes.
On trouve de nombreuses espèces de ptéridophytes fossiles, ce qui est une preuve de leur épanouissement pendant les périodes du Carbonifère et du Permien (les restes forment le charbon). Des groupes importants comme les Equisétinées étaient arborescentes.
On trouve quatre classes de ptéridophytes : les Psilophytinées, les Lycopodinées, les Equisétinées et les Filicinées.
II La sous-classe des Filicinées, avec Dryopteris filixmas.
Elle est aussi appelée «la fougère mâle ».
A Le sporophyte ou l’appareil végétatif.
1 L’appareil végétatif.
C’est une plante feuillée constituée, par une tige souterraine (rhizome) portant un bouquet de grandes feuilles pennatiséquées à son extrémité supérieure, par un pétiole, un rachis (l’axe central) et par des feuilles (pennes) divisées en pinnules. Les feuilles ont une préfloraison circinée (ou en crosse).
2 Les racines.
Elles sont adventives, avec une stèle (ou cylindre central) à deux pôles ligneux à différenciation centripète ainsi que deux pôles libériens.
3 Le rhizome.
C’est le cylindre central. Il est composé d’un épiderme, d’un sous-épiderme (en assises de cellules), de parenchymes à réserves amylacées (avec de l’amidon), et de deux cercles concentriques de stèles. Le cercle externe donne les traces foliaires qui donneront les feuilles. Le cercle interne permet la vascularisation propre du rhizome. Chaque stèle a son endoderme à bande de Caspary et son péricycle.
4 Les feuilles : sont chlorophylliennes avec des stomates.
5 Les sporanges.
Ils sont à la face inférieure des pinnules. Les sporanges se développent en amas (ou sores) constitués par une colonne centrale sur laquelle sont fixés les sporanges protégés par une membrane (c’est l'indusie). Le sporange n’a pour origine qu’une seule cellule superficielle. La division transversale sonne une cellule interne qui va former le pédicelle et une cellule externe qui donne un sporange de type leptosporangié. La tête du sporange est constituée par un archéspore et des cellules pariétales (autour de la première) puis elles se divisent. On obtient deux assises de cellules. L’assise externe donne le tapis qui est composé de cellules nourricières et l’assise interne qui donnera les cellules mères des tétraspores. Les cellules pariétales donnent la paroi du sporange constitué par un arc de cellules à paroi épaissie sur trois côtés, ainsi que des cellules normales qui forment le stromium (zone de moindre résistance qui s’ouvre par temps sec). Le tapis sert de nourriture aux cellules mères des tétraspores.
6 La réduction chromatique, formation des spores.
Les cellules mères subissent la réduction chromatique et donne, chacune 64 spores à N chromosomes. Les spores sont entourées d’une membrane épaisse formée de trois couches. Les spores sont libérées par l’ouverture du sporange et germent par temps humide. Ils donneront un gamétophyte ou prothalle.
B Le gamétophyte, à N chromosomes.
1 Son appareil végétatif.
Les spores donnent des filaments de 5 ou 6 cellules portant des rhizoïdes incolores : c’est le prothalle. Quand il se développe, il donne une lame aplatie et cordiforme (en forme de cœur). Cette lame est composée d’une seule assise cellulaire sauf dans le coussinet (la région médiane) portant les rhizoïdes. Le prothalle est chlorophyllien, autotrophe et autonome.
2 Les anthéridies.
Sur la partie inférieure du prothalle, il y a apparition des nombreuses premières anthéridies. Chaque cellule à pour origine une cellule épidermique. Les anthéridies sont constituées d’une paroi (4 ou 5 cellules) et par un couvercle (2 ou 3 cellules). A l’intérieur, les cellules donnent 32 anthérozoïdes rubanés avec un bouquet de flagelles et des vésicules qui se résorbent, puis ces anthérozoïdes sont libérés par l’ouverture du couvercle.
3 L’archégone.
Elle apparaît plus tardivement, à partir d’une cellule superficielle. Celle-ci se trouve à la face inférieure du prothalle (à terre, comme le «mâle »). Sous le col, composé de 7 ou 8 cellules, se trouvent l’oosphère et des cellules du canal du col. L’espèce est monoïque et homothallique.
4 La fécondation.
L’apex de la cellule s’ouvre et les cellules du col se gélifient en donnant un mucilage. Les cellules mâles sont attirées par chimiotachtisme et pénètrent et vont féconder l’oosphère. Ceci donnera un zygote à 2N. C’est une zoïdogamie (ce qui est différent d’un oogamie).
5 Le retour du sporophyte
Le zygote se transforme en embryon qui développe une racine, une tige et une feuille, plus un pied suçoir qui se fixe (pas longtemps) sur le thalle. Il gagne rapidement son autonomie.
C Conclusion.
Le sporophyte devient prépondérant, autonome. Le cycle est haplodiplophasique, digénétique. Il y a zoïdogamie avec un milieu extérieur liquide. L’espèce est isosporée (même taille des spores) et homothallique.
III La sous-classe des lycopodinées, avec Selaginella sp.
A Le sporophyte.
1 L’appareil végétatif.
C’est une plante feuillée de type herbacé. Ses tiges sont vascularisées, se ramifient de manière dichotomique et portent de petites feuilles (les lancéoles) en disposition spiralée ou opposée. Sur la face supérieure, près de l’insertion de la tige, on a quatre ligules (des petites languettes). Les racines ne sont pas directement fixées sur la tige mais sur des axes rhizophores.
2 Le sporange.
Il se développe à l’aisselle de feuilles dites fertiles : les sporophylles. Elles sont groupées en épis sporangifères. On sommet des rameaux, on trouve les strobiles.
On trouve deux types de sporanges sur le même pied :
· Les mégasporanges qui donnent quatre tétraspores volumineuses.
· Les microsporanges qui donnent de nombreuses microspores.
Les épis sporangifères portent les mégasporanges à leur base et les microsporanges à leur apex.
Remarque : on les a parfois sur deux rangs verticaux chacun.
Dès les premiers stades, le développement des méga et microsporanges sont identiques. Ils ont : une paroi composée de plusieurs cellules, de cellules nourricières en manchon qui vont donner le tapis et les cellules mères des tétraspores.
Le développement est eusporangié (à partir d’un petit groupe de cellules superficielles).
3 La réduction chromatique.
Dans les microsporanges, les cellules mères subissent la réduction chromatique et donnent de nombreuses microspores.
Dans les mégasporanges, toutes les cellules mères avortent, sauf une, qui va subir la réduction chromatique et donnera quatre tétraspores volumineuses.
B Les prothalles.
1 Le gamétophyte mâle, ou, le microgamétophyte.
Une fois que les microspores sont libérées des microsporanges, on obtient deux prothalles dont un qui est à l’origine des anthéridies (l’anthéridie, après de multiples divisions, est formée d’une paroi entourant quatre cellules spermatogènes : les gamétogènes). Les cellules spermatogènes donnent plusieurs anthérozoïdes biflagellés, libérés après la rupture de la spore.
2 Le gamétophyte femelle.
Son développement débute dans le mégasporange. Le noyau se divise plusieurs fois et donne un prothalle coenocytique (un noyau dans une cellule), puis il y a apparition de cellules à l’un des pôles.
Le prothalle adulte est chlorophyllien au niveau de l’ouverture et porte des rhizoïdes. Du côté opposé à l’ouverture, on a une zone riche en amidon. La partie supérieure des archégones réduits est constituée d’un col de 2 ou 3 cellules (de chaque côté). L’oosphère est entourée par des cellules prothalliennes et par deux cellules du canal du col.
L’espèce est dioïque.
3 La fécondation.
Les anthérozoïdes libérés, nagent dans l’eau vers les archégones (ils sont attirés par chimiotachtisme). Il y a alors, fécondation d’une ou plusieurs oosphères, qui entraînera le développement d'un zygote. La fécondation est une zoïdogamie en milieu extérieur liquide.
4 Le développement du nouveau sporophyte à 2N.
L’œuf se divise en deux et donne alors une cellule supérieure (suspenseur) et une cellule inférieure qui, en se divisant, va donner un embryon avec une tige, une racine et un pied, fixant au départ, l’embryon sur le prothalle.
L’embryon et la jeune plantule restent un peu fixés au prothalle puis deviennent autonomes.
On a un cycle avec deux types de spores : c’est une hétérosporie. Ce phénomène entraîne la formation de deux types de gamétophytes, d’où l’hétérothallisme.
Il y a réduction du gamétophyte et de ses organes reproducteurs.
C’est une espèce haplodiplophasique, digénétique dont les spores sont une forme de résistance et de dissémination de l’espèce.
Chapitre 7 : Les phanérogames ou spermaphytes : les Gymnospermes.
Ce sont des végétaux vasculaires dont la fécondation est indépendante du milieu extérieur et dont les éléments reproducteurs (gamétophytes) sont les grains de pollen chez les mâles et les ovules chez les femelles.
On a deux sous-ensembles majeurs : les gymnospermes dont les ovules et les graines sont nus, et les angiospermes chez qui la graine est dans un fruit.
Ils apparaissent au Dévonien. Leur épanouissement à lieu à la fin de l’ère primaire et pendant l’ère secondaire. Maintenant, ce groupe est en déclin. Les plus nombreux sont les coniférales. Ce sont surtout des arbres et arbustes à feuilles persistantes.
Leur anatomie :
- trachéïdes à ponctuations aréolées.
- le bois secondaire est homoxylé.
- ils possèdent de nombreux canaux à résine (d’où le terme de résineux).
On trouve trois classes : les Cycadophytes, les Coniférophytes, les Gnétophytes. On a toutefois un autre type de classification avec quatre subdivisions : les Cycophyta, les Ginkgophyta, les Coniférophyta, les Gnétophyta.
I Le Ginkgo biloba de la division des Ginkgophyta.
C’est un arbre asiatique qui a été introduit en France au 18éme siècle.
A Le sporophyte à 2N
1 L’appareil végétatif.
C’est une espèce dioïque (mâle ou femelle). Ses feuilles sont caduques avec deux sortes de rameaux. On a les pousses longues, à entre-nœuds allongés et à croissance indéfinie : ce sont les auxiblastes. On a ensuite les pousses courtes, qui sont sexuées et à croissance définie : ce sont les mésoblastes.
2 L’appareil reproducteur mâle (à fleurs mâles).
Les fleurs sont constituées par des bouquets d’étamines situés à l’aisselle des feuilles. Les étamines sont formées par un filet portant deux sacs polliniques. A maturité, il y a ouverture du sac par la fente longitudinale : c’est la libération du pollen.
Les sacs polliniques proviennent de la différenciation d’un massif d’archéspores (à 2N) se divisant et donnant une assise nourricière (le tapis) et les cellules mères des microspores à 2N.
3 L’appareil reproducteur femelle (à fleurs femelles).
Ces fleurs sont situées à l’extrémité des rameaux courts. Elles sont sur un axe (le pédicelle), qui porte deux ovules, dont une qui avortera en formant une cupule. Les ovules (ou, masses cellulaires, ou nucelle) sont entourés par un tégument interrompu, au sommet, par le micropyle.
B Les gamétophytes.
1 Le gamétophyte mâle.
Il est dans le sac pollinique. Les archéspores à 2N subissent la réduction chromatique pour donner des microspores qui vont évoluer en grains de pollen. Un grain de pollen est composé de deux cellules prothalliennes protégées par l’exine, d’une cellule gamétogène, d’une grande cellule végétative protégée par l’intine. La partie végétative du grain de pollen est constituée par une cellule prothallienne et par les deux autres cellules non prothalliennes.
2 Le gamétophyte femelle.
Dans le nucelle, l’archéspore donne une cellule mère qui subit la réduction chromatique et qui donne 4 mégaspores (seule la plus interne va se développer en donnant le gamétophyte femelle : l’endosperme). Cet endosperme va subir de nombreuses divisions et va être formé de deux parties : par un endosperme coenocytique (avec beaucoup de noyaux) et par un endosperme définitif qui est un ensemble de cellules où s’accumulent des réserves (amidon et lipides). Au sommet de l’endosperme, deux archégones se différencient.
C Pollinisation et germination.
Le pollen est disséminé par le vent au printemps alors que les ovules ne sont pas encore mûres. Il y a alors une sécrétion de liquide mucilagineux par le micropyle qui permet la fixation des grains de pollen. Il y résorption de l’ensemble, liquide plus pollen, vers la chambre pollinique qui se referme. Le grain de pollen va germer pour donner un microprothalle, en se fixant dans le nucelle. La cellule gamétogène se divise et donne une cellule du pied (ou cellule du tube) et une cellule du corps (reproductrice). Cette cellule reproductrice donne deux anthérozoïdes (ou spermatozoïdes flagellés).
D Fécondation et formation de l’œuf et du sporophyte.
Libérés du grain de pollen, les spermatozoïdes nagent dans le liquide du nucelle et l’un d’eux va féconder l’oosphère (c’est une zoïdogamie à milieu interne à l’organisme liquide). Cette fécondation est suivie de la formation de zygote (ou œuf). Le noyau du zygote se divise immédiatement et donne un pré-embryon qui se différencie en deux zones. Une zone supérieure qui est un suspenseur (c’est aussi le col de l’archégone), et une zone inférieure méristématique : l’embryon qui, complètement différencié, est formé d’une radicule, d’une tigelle, d’une gémule et de deux cotylédons.
E Le développement de l’ovule fécondé.
A maturité, l’ovule donne une pré-graine dont le tégument se divise en trois couches. Le nucelle persiste en une mince couche. L’endosperme occupe toute la partie centrale. Les réserves sont dans l’endosperme avant la fécondation. Les ovules peuvent tomber avant d’être fécondés. Il n’y a pas de vie ralentie avant la germination.
Chez le Ginkgo, on assiste à la première apparition d’un ovule (mégasporange) nu. Le gamétophyte n’a pas de vie autonome : le gamétophyte mâle est parasite du sporophyte et le gamétophyte femelle est réduit.
La fécondation est une zoïdogamie avec le liquide fourni par l’ovule. Le cycle est haplodiplophasique, il est surtout diplophasique car c’est la seule forme de vie autonome.
II Le pin coniferophyta.
Ce sont des arbres de grande taille dont les feuilles sont groupées par deux en aiguilles. L’espèce est monoïque. L’appareil végétatif constitue le sporophyte. Cette espèce entraîne une acidification du sol qui devient alors moins fertile.
A Les appareils reproducteurs (sur le sporophyte).
1 L’appareil mâle.
Il est constitué de cônes groupés en épis à la base de jeunes pousses. Les cônes sont des associations, autour d’un axe de feuilles modifiées (ou écailles), qui forment les étamines. Le sporange (le sac pollinique) est situé à la face inférieure des écailles. Le pollen est libéré par une ouverture longitudinale. Chaque grain est limité par une paroi riche en sporopollénine, dilatée latéralement en bâtonnets, ce qui favorise la dissémination par éochorie. Une microspore haploïde donne un grain de pollen. Le gamétophyte mâle se développe dans le sac pollinique, puis, à maturité, le grain est constitué de deux cellules prothalliennes, d’une cellule reproductrice et d’une cellule végétative. Le grain est disséminé pendant sa phase de vie ralentie.
2 L’appareil femelle.
Des cônes se développent au sommet des jeunes pousses et sont une association d’écailles à l’aisselle des bractées. Au printemps de la première année, deux ovules sont à la face supérieure de chaque écaille. Les ovules ne sont pas protégés par une formation sporophytique, ce qui les laisse nus. Le nucelle est entouré d’un tégument laissant une ouverture apicale (le micropyle).
Quand l’archéspore augmente de taille, elle subit la méiose qui donne quatre mégaspores dont seule, la plus profonde, persiste. Le noyau se divise plusieurs fois et donne le gamétophyte femelle. L’endosperme coenocytique arrête vite de se développer.
B La pollinisation.
Elle est assurée par le vent. Les grains de pollen sont captés par le mucilage et arrivent au contact du nucelle. Ils germent alors en produisant un tube pollinique dans lequel s’engage le noyau de la cellule végétative. Puis la cellule reproductrice se divise en deux cellules dont une cellule spermatogène (anthéridie). Le développement s’interrompt alors jusqu’au printemps suivant.
C La fécondation.
Le développement du gamétophyte femelle reprend car il y a formation de cellule dans l’endosperme. Quand celle-ci a terminé, il y a individualisation de 2 ou 3 archégones. Les archéspores donnent une oosphère volumineuse à col réduit.
Le pollen à une reprise de métabolisme plus tardive (seulement quelques jours avant la fécondation). L’anthéridie se divise en deux gamètes mâles non limités par une paroi.
Quand le tube pollinique, dans le col de l’archégone, déverse son contenu dans l’oosphère, on a une cellule mâle qui dégénère et une qui s’unit à l’oosphère pour former un zygote à 2N. Le gamète mâle n’a jamais été mobile : c’est une fécondation par siphonogamie.
D Le zygote.
Après s’être divisé deux fois, le zygote donne un embryon coenocytique à quatre noyaux dans la partie basale (à l’opposé du col de l’archégone).
Les cellules les plus basales sont les cellules embryonnaires. Elles sont surmontées par la cellule du suspenseur qui s’allonge de manière importante. Cette croissance donne une séparation de quatre cellules embryonnaires qui donnent quatre embryons dont trois qui dégénèrent. L’embryon qui reste s’organise en une jeune plantule.
E La graine.
Pendant le développement de l’embryon, les cellules de l’endosperme et des jeunes feuilles de la plantule (cotylédons) se chargent de réserves ; le tégument s’épaissit, se lignifie ; l’endosperme et la plantule se déshydratent et sont accompagnés d’une réduction des activités métaboliques. C’est le passage à la vie ralentie. L’ovule est transformé en une graine pourvue d’une aile. Après une période de vie ralentie, quand les conditions sont favorables, la graine germe et donne une nouvelle plante feuillée.
Le cycle est caractérisé par l’alternance d’une génération sporophytique dominante et d’une génération gamétophytique réduite. Cette dernière se développe au sein du sporophyte, en parasite de celui-ci. Les différences essentielles avec le Ginkgo portent sur la fécondation (ici on à siphonogamie) et sur la graine qui, ici, est une véritable graine capable de vivre à l’état de vie ralentie.
Chapitre 8 : Les Angiospermes.
I Généralités.
C’est un sous-embranchement des spermaphytes qui est divisé en deux sous-classes : les monocotylédones et les dicotylédones. Ces angiospermes sont caractérisés par la présence de l’ovule dans un ovaire et de la graine dans un fruit. Les organes reproducteurs (à l’origine des gamètes) sont placés dans une structure particulière : la fleur.
II Cas d’un dicotylédone avec Ranunculus acris.
A L’appareil reproducteur mâle (les étamines)
Les étamines sont constituées d’un filet (pédicelle rattaché au réceptacle floral) et d’une anthère où se différencie le pollen. Chaque anthère est constituée de deux loges, symétriques par rapport au filet, qui comportent chacune deux sacs polliniques. Dans une étamine jeune, chaque étamine jeune comprend une masse centrale de grosses cellules à noyaux volumineux : ce sont les archéspores. Elles sont entourées de plusieurs assises de cellules : par les cellules nourricières qui forment le tapis, par plusieurs assises transitoires, par une assise mécanique dont les cellules lignifiées interviennent dans l’ouverture de l’anthère et par un épiderme.
Les archéspores subissent la réduction chromatique et donnent, chacun, quatre microspores (à N chromosomes).
Remarque :Chez les monocotylédones, il y a cloisonnement après chaque division cellulaire, alors que chez les dicotylédones, il y a cloisonnement toutes les deux divisions.
Les microspores se divisent une nouvelle fois et s’entourent d’une paroi épaisse, ornementée, et constituée de deux couches (l’exine et l’intine). L’exine est percée de pores germinatifs. A maturité, chaque grain de pollen contient une cellule végétative à gros noyau et une cellule reproductrice à petit noyau. Le grain de pollen est le gamétophyte mâle qui va se disséminer. Ce grain a commencé son développement dans l’anthère, avant de se charger en matériaux de réserve et de subir la déshydratation qui le fait passer à l’état de vie ralentie (parfois, on a seulement des loges polliniques). L’ouverture des anthères se fait grâce à la dessiccation. La longévité d’un grain de pollen est variable (graminées : un jour ; rosacées : 100 jours).
B L’appareil reproducteur femelle.
Il est constitué par :
- Les carpelles qui sont, chacun, formés par une partie renflée et creuse qui contient un ovule.
- Un style court, terminé par un stigmate formé de papilles.
- L’ovule est constitué par une partie tégumentaire formée de deux membranes. Ces deux membranes sont interrompues par une minuscule pore : Le micropyle. L’ovule est inséré sur la partie renflée de l’ovaire, sur le placenta. Cet ovule est relié au funicule.
Chez la renoncule, l’ovule est anatrope : le micropyle est en bas, proche du placenta. Le plan de raccordement du funicule et de l’ovule s’appelle le hile. (anatrope = complètement renversé).
Dans le funicule, pénètre un faisceau de tissus conducteurs qui se ramifient au niveau de la chalaze. A l’intérieur de l’ovule se trouve le nucelle.
Remarque :On a différents types d’ovules : droits (ou orthotropes), les axes du funicule et du nucelle sont confondus ; courbes (ou campylotropes), les axes du funicule et du nucelle sont perpendiculaires.
C La réduction chromatique.
Le sac embryonnaire, dans le nucelle, représente le prothalle femelle. A l’origine, c’est une cellule épidermique qui s’est divisée en une cellule pariétale (qui avorte) et en une cellule sporogène. C’est cette dernière qui va subir la réduction chromatique en donnant quatre tétraspores dont trois qui vont dégénérer. La spore la plus interne va subsister et va constituer le sac embryonnaire.
D Le gamétophyte femelle.
Après les divisions du mégaspore, les noyaux se répartissent en trois complexes :
- Le pôle microplylaire : l’oosphère est associée à deux synergides.
- Le pôle de la chalaze : c’est un groupe de trois cellules, les antipodes
- La zone centrale qui contient deux noyaux «polaires ».
E Pollinisation et germination.
Pour la renoncule, la pollinisation se fait par les insectes et par le vent. Le grain de pollen va se déposer sur le stigmate, va germer en produisant un long tube pollinique qui s’enfonce dans le nucelle jusqu’à l’ovule. Le noyau reproducteur du pollen se divise en deux spermatozoïdes.
Le tube peut pénétrer par le micropyle : c’est une porogamie.
Il peut rentrer par la chalaze : c’est une chalazogamie.
La durée de croissance du tube va de quelques heures à quelques mois.
1 La fécondation.
Le contenu du tube se décharge dans le sac embryonnaire. L’un des spermatozoïdes s’unit avec l’oosphère et donne le zygote principal (à 2N). L’autre spermatozoïde s’associe aux noyaux polaires et va former un zygote accessoire (à 3N). C’est la double fécondation des angiospermes.
2 La graine.
Le zygote principal donne une plantule avec deux feuilles embryonnaires (les cotylédons). Le zygote triploïde va donner l’albumen. Au cours du développement de la plantule, les réserves s’accumulent dans l’albumen et dans les cotylédons. Les téguments se lignifient (durcissement). Passage à l’état de vie ralentie, puis, dispersion.
On a différents types de téguments :
- La caroncule, qui est un bourgeonnement au niveau du micropyle du ricin.
- L’arille, qui est un bourgeonnement au niveau du hile.
- L’arillade qui est un revêtement recouvrant la graine.
3 Les différents types de graines.
Selon l’évolution du nucelle, on obtient :
4 La germination.
Elle a lieu quand la graine est dans un bon état physiologique et quand les conditions du milieu sont favorables. Un bon état physiologique est caractérisé par un grand pouvoir germinatif qui dépend de l’état de maturité des graines (certaines germent immédiatement mais pendant un temps très court, d’autres ont une germination différée : elles ne peuvent germer qu’après une «dormance » ayant plusieurs origines : les téguments de l’embryon sont immatures, des inhibiteurs de levée de dormance sont présents et doivent être éliminés par des conditions favorables du milieu : réhydratation).
Cette germination a lieu quand des conditions favorables comme la concentration en oxygène, la température, la quantité de lumière, ou, des facteurs biologiques sont réunis. Il y a alors croissance de la radicule et émission d’un bourgeon (la gemmule) qui donnera les premières feuilles. Il y a parfois une tigelle entre la radicule et la gemmule.
On a deux types de germination :
· La germination épigée (exemple, le haricot). La graine est soulevée hors du sol car il y a un accroissement rapide de la tigelle qui donne l’axe hypocotyl qui soulève les deux cotylédons hors du sol. La gemmule se développe (après la radicule) et donne une tige feuillée au-dessus des deux cotylédons. Le premier entre-nœud donne l’épicotyl. Les premières feuilles, au-dessus des cotylédons sont les feuilles primordiales (elles sont plus simples que les futures feuilles).
· La germination hypogée (exemple, le maïs). La graine reste dans le sol, la tigelle ne se développe pas et les cotylédons restent dans le sol.
Bilan sur les angiospermes :
Ils ont un cycle haplo-diplopasique (surtout sporophytique). Les gamètes femelles sont parasites du sporophyte. La fécondation est une siphonogamie (attention, c’est une double fécondation).
III Les archégoniates cormophytes.
A partir des mousses, l’évolution des gamètes mâles et femelles donne une diminution du territoire végétatif du prothalle et la perte de son indépendance.
Passage au milieu aérien et mode de vie.
Le gamétophyte mâle se développe à partir des réserves accumulées dans les spores. Il y a diminution du nombre d’anthéridies.
Le gamétophyte femelle : le prothalle cesse d’accumuler des réserves, elles seront élaborées après la fécondation. Ce sont les gamétophytes les plus évolués.
La réduction des gamétophytes et des gamétanges est une stratégie évolutive. Cette stratégie entraîne, un allongement du cycle de développement (80 heures pour les champignons et plusieurs années pour les fleurs), la suppression partielle de la génération haploïde indépendante (fragile), l’affranchissement des aléas d’un approvisionnement énergétique et en eau, en permettant le développement de prothalles parasites du sporophyte.
L’évolution a d’abord formé des gamètes mobiles dans l’eau pour les algues et les mousses, puis des gamètes véhiculés, pour les archégoniates, par les tubes polliniques grâce à la siphonogamie. Celle-ci permet un affranchissement de l’eau pour effectuer la fécondation. C’est aussi le passage d’une fécondation simple à une fécondation double qui conduit chez les angiospermes à la formation d’un embryon « nourrice » ou albumen.
Sommaire du cours de
Cormophytes.
|
Partie 1 : Diversité des cormophytes (bryophytes, ptéridophytes, spermaphytes).
|
|
I Evolution des végétaux.
II Evolution de l’appareil reproducteur.
III Les bases de la classification (classe, ordre, famille, genre, espèce).
IV Les bryophytes : classification, écologie, utilisation.
V Les ptéridophytes : classification, écologie, utilisation.
VI Les gymnospermes.
VII Les angiospermes. |
I Introduction.
II Les rythmes biologiques.
III Les types écologiques.
IV Multiplication et dissémination sous forme végétative.
V Reproduction sexuée et stratégie de pollinisation.
IV Conservation et dissémination des graines.
V La fixation des plantes. |
Partie 1 :
Diversité des cormophytes
(bryophytes, ptéridophytes, spermaphytes).
Le cormus est l’appareil végétatif, constitué d’une tige différenciée.
Les végétaux apparaissent il y a 2 milliards d’années. Les premiers « végétaux » sont les cyanophycées (chlorophylliennes). Grâce à la photosynthèse, elles permettent de faire diminuer la concentration en CO2 et d’augmenter la concentration en O2.
Il y aurait un milliard d’années seraient apparus les eucaryotes. On explique cette apparition par la théorie de l’endosymbiose : procaryote pris par un eucaryote. Les végétaux deviennent pluricellulaires, mais toujours marins.
La conquête du milieu terrestre a eu lieu vers –600 millions d’années. Il y a alors obligation de s’adapter : modifications des organes reproducteurs et apparitions de vaisseaux conducteurs. Peu à peu, apparaissent alors des plantes vasculaires (ptéridophytes : cryptogames vasculaires).
Le développement des fougères se fait surtout durant l’ère primaire. Elles seront remplacées, durant le secondaire, par les gymnospermes.
Il y a 130 millions d’années apparaissent les angiospermes (graine enveloppée). De –130 à –90 millions d’années, on assiste à un big-bang des angiospermes. A la fin du crétacé, c’est la domination des angiospermes qui dure jusqu’à l’actuel. Le succès des angiospermes est du aux facultés d’adaptation, de migration…
I Evolution des végétaux.
A Les thallophytes.
Des algues unicellulaires, on passe aux algues pluricellulaires avec apparition de thalle. Il n’y a pas d’organe bien différencié sauf chez les laminaires qui ont des organes rudimentaires avec spécialisation d’organes reproducteurs.
B Les bryophytes.
Chez les bryophytes apparaissent des organes permettant la conquête du milieu terrestre : folioles et tige rudimentaire. Ce sont les premiers cormophytes, bien que leur cas soit litigieux car ils n’ont pas de racine mais des rhizoïdes. Il existe encore des bryophytes à structure avec thalle.
C Les ptéridophytes.
Les ptéridophytes ont de vraies tiges avec des vaisseaux conducteurs de sève (? trachéophytes). Ce type de végétaux est apparu dès le dévonien. Il y avait des forêts constituées de Rhynia (disparus aujourd’hui). Ces derniers ont des crampons mais pas de racine. Il existait des fungi symbiotiques avec les Rhynia (? endomycorhizes).
Le Psilotum ressemble aux Rhynia : ils présentent toujours un appareil conducteur mais avec une tige simplifiée : le télome.
Les Lépidodendrons constituèrent des forêts. Ils pouvaient atteindre une hauteur de 40 mètres. Ils ont eu une apogée au carbonifère.
Ce groupe présente une grande diversité de l’appareil végétatif.
D Les gymnospermes.
Les gymnospermes remplacèrent les ptéridophytes à la fin du primaire. Ils sont uniquement ligneux, avec un développement de xylème secondaire ( « bois »). Au niveau du xylème, on trouve des trachéides qui ont un rôle de transport et de soutien.
« bois »). Au niveau du xylème, on trouve des trachéides qui ont un rôle de transport et de soutien.
E Les angiospermes.
Les angiospermes ont un développement maximal entre –130et –90 millions d’années. Il y a apparition de vaisseaux dont le rôle est uniquement de conduire les sèves. Il y a un partage des tâches (vaisseaux + cellules à lignine).
II Evolution de l’appareil reproducteur.
La méiose donne des spores haploïdes qui ne sont pas directement des gamètes. Les spores subissent donc une phase de développement et de croissance. Une fois apparu, le gamétophyte produira des gamètes.
Il y a donc développement d’un gamétophyte qui est haploïde. Il est issu d’une méiospore et va produire les gamètes.
Les méiospores viennent du sporophyte qui est diploïde et qui, en subissant la méiose, va donner les méiospores.
On trouve une part respective au sporophyte et une respective au gamétophyte.
A Les bryophytes.
La partie visible, chlorophyllienne est un gamétophyte (à N chromosomes).
Les spores donnent un protonéma (?thalle) à courte durée de vie où se développent une tige avec des rhizoïdes (= gamétophyte).
Il y a développement de gamétanges sur le gamétophyte : ce sont les organes qui produisent les gamètes. Les gamétanges mâles donnent les anthéridies alors que les gamétanges femelles donnent les archégones. Selon les cas, les organes mâles et femelles sont portés par des pieds différents ou bien par le même pied. Les anthérozoïdes sont les gamètes mâles, ciliés. L’oosphère est le gamète femelle.
L’anthérozoïde est porté par une goutte d’eau, attiré par chimiotactisme. Il y a ensuite rencontre puis fusion des gamètes au sein de l’archégone : le développement du sporophyte est constitué par l’ensemble : soie + capsule + coiffe.
Ce sporophyte donne alors des méiospores et le cycle peut recommencer.
C’est un cycle digénétique, haplodiplophasique avec une dominance de la phase haploïde gamétophytique.
B Les ptéridophytes.
Visuellement, on observe le sporophyte (à 2N). Sur la face inférieure des frondes, on a des amas de sporanges. Ces derniers donneront les spores qui eux, donneront un gamétophyte de quelques millimètres. Le gamétophyte donne des gamétanges mâles et femelles. Les gamétanges mâles forment des anthérozoïdes ciliés qui iront féconder l’oosphère. Il y a apparition d’un zygote qui donne un sporophyte. Ce dernier se développe sur le gamétophyte.
C’est un cycle digénétique, diplohaplophasique, avec dominance de la phase sporophytique (homosporie et homo-prothallie).
Chez les Sélaginelles, il y a une évolution supplémentaire. A l’aisselle de certaines feuilles, on peut voir des organes reproducteurs : les hétérosporophylles.
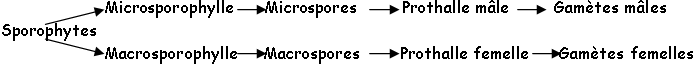
Les gamètes mâles (anthérozoïdes) et femelles vont fusionner (fécondation) et redonner un sporophyte. Le prothalle femelle reste au niveau de au niveau du sporophyte (est donc dépendant) : C’est l’endoprothallie.
C Les gymnospermes.
Les gymnospermes sont des arbres à 2N chromosomes. Ils présentent des macrosporophylles ou des microsporophylles. L’appareil femelle est au niveau des « pommes de pin » ; l’appareil mâle est au niveau des organes mâles.
Les macrosporanges sont formés par une enveloppe qui entoure la macrospore ? l’ovule. La macrospore va se diviser de nombreuses fois et donner l’endosperme où se forment les archégones et les oosphères (le prothalle femelle est l’endosperme).
Les microsporophylles donnent des microsporanges (sacs polliniques). Ils vont subir la réduction chromatique et donner des grains de pollen (= prothalle mâle).
La fécondation est réalisée avec élaboration d’un tube pollinique qui permet un affranchissement du milieu aqueux. Il va y avoir fusion des noyaux puis formation de la graine.
D Les pré-spermaphytes, exemple de Ginkgo biloba.
Le Ginkgo est appelé « fossile vivant » ; c’est une espèce dioïque.
La fécondation a lieu après la séparation de l’ovule avec la plante mère. L’ovule et l’endosperme se développent sans qu’il y ait obligatoirement fécondation.
Les anthérozoïdes sont ciliés. Ils entrent en contact avec l’ovule et pénètrent dans la chambre pollinique. La fécondation peut avoir lieu dès que l’ovule est tombé par terre.
Après la fécondation, il n’y a pas de dormance, mais développement.
On trouve encore des anthérozoïdes ciliés et on note l’absence de tube pollinique.
E Les angiospermes.
Chez les angiospermes, la plante est le sporophyte à 2N. Il va produire des spores qui vont se développer en microsporophylles (étamines) ou en macrosporophylles.
Les étamines donnent des microsporanges (sacs polliniques) qui libèrent des microspores. Les macrosporophylles donneront un ovule entouré d’un ovaire ou d’un carpelle (l’appareil reproducteur).
Après une série de divisions, suivies de la méiose, le gamétophyte mâle (ou prothalle) donne les grains de pollen (ce sont aussi le gamétophyte). Ce gamétophyte est très réduit. La paroi est constituée de deux parties : l’exine et l’intime. L’exine est résistante et forme des ornementations qui permettent de déterminer les genres et familles de végétaux. A l’intérieur, on trouve une cellule spermatogène et une cellule végétative.
Au moment de la pollinisation, la cellule végétative donnera le tube pollinique et la cellule spermatogène donnera deux noyaux reproducteurs (deux spermatozoïdes).
Chez l’appareil femelle, le carpelle se met en place avec une macrospore qui se divise en sacs embryonnaires qui constituent le prothalle femelle (le gamétophyte femelle). Ce prothalle reste inclut dans le sporophyte. Dans la partie supérieure, on trouve trois cellules : les antipodes. Au milieu, on trouve deux noyaux polaires. A la base, on a l’oosphère et deux synergides (deux cellules proches).
Au moment de la fécondation, quand le tube pollinique atteint le sac embryonnaire, il y a double fécondation. Les deux anthérozoïdes pénètrent dans le sac embryonnaire. L’un d’eux va fusionner avec l’oosphère et donner l’œuf embryon. Le second, fusionne avec les deux noyaux polaires et donne une cellule à 3N.
C’est l’œuf albumen.
La double fécondation est caractéristique des angiospermes. Elle permet l’obtention d’un embryon et de réserves, synthétisées par l’œuf.
L’ovule va évoluer en téguments (durcissement des téguments de la graine) et le sac embryonnaire en embryon et albumen.
L’ovule des végétaux est différent de l’ovule des animaux.
III Les bases de la classification (classe, ordre, famille, genre, espèce).
La classification vient de la nomenclature binomiale, officialisée par un suédois. Il a mis en place un système de classification, différent pour les végétaux et pour les animaux :
Pour les végétaux, il y a prise en compte des appareils reproducteurs (surtout des étamines). ? Ce système est artificiel car on regroupe des espèces qui ne se ressemblent pas au niveau végétatif.
Par exemple, les plantes grasses peuvent appartenir à des familles très diverses car leur appareil sexuel est distinct même si elles possèdent un appareil végétatif très comparable.
IV Les bryophytes : classification, écologie, utilisation.
Les bryophytes sont divisées en deux groupes :
- Les hépatiques. Ces plantes présentent une sorte de lame foliacée ou une tige feuillée à symétrie bilatérale.
- Les mousses. La tige est foliée, à symétrie axiale. Les sporogones sont bien développés.
A Les Hépatiques.
1 Hépatiques à tige feuillée. (Ordre des Jungermanniales).
On observe une symétrie bilatérale car les feuilles sont incluses dans un plan. Elles vivent dans les milieux humides (environ 7500 espèces).
2 Hépatiques à thalle. (Ordre des Marchantiales).
Exemple de Marchantia polymorpha.
Cette plante vit au bord des ruisseaux, des fontaines. L’appareil végétatif est constitué d’une tige foliacée. C’est une des premières plantes à recoloniser les sites incendiés. Elle peut se reproduire, soit par multiplication végétative, soit par reproduction sexuée.
- La multiplication végétative. Il y a élaboration de corbeilles à propagules (avec des morceaux de thalle qui seront disséminés par le vent ou par l’eau).
- La reproduction sexuée. Le thalle va former des gamétophytes qui donnent des gamétanges (les « parapluies »). Les gamétanges mâles donnent des anthéridies qui vont féconder les archégones produits par les gamétanges femelles. Cette fécondation donne un sporophyte à 2N qui reste sous le « parapluie ».
B Les Mousses.
Les mousses présentent une symétrie axiale. Elles sont regroupées en deux ordres principaux : les sphagnales avec deux sortes de feuilles et les bryales avec une seule sorte de feuilles.
1 Les Sphagnales.
On ne trouve qu’une famille, les Sphagnaceae, et un seul genre, Sphagnum (sphaigne). Les espèces sont environ 300 et 35 sont présentes en France. Ce sont des plantes typiques des tourbières.
Ce sont les Sphagnum qui sont à l’origine de la tourbe.
Les tourbières sont un biotope avec des eaux stagnantes. Le milieu montre une minéralisation extrêmement lente de la matière organique. Très peu de végétaux peuvent coloniser ces milieux. Les sphaignes ont une croissance apicale. La partie inférieure de la tige se dessèche, meurt, mais reste présente et s’accumule. ? Il y a un épaississement de la tourbe au fil du temps. Les tourbières sont généralement présentes dans les milieux de hautes latitudes. Les tourbières sont riches en azote et le pH des eaux est acide.
Dans les tissus des Sphaignes, on distingue deux types cellulaires :
- Des cellules allongées qui vont donner un réseau chlorophyllien.
- Des cellules particulières, les hyalocystes (ou hydrocystes), qui sont de grandes cellules mortes rigidifiées par un épaississement. Elles ont la capacité de se remplir d’eau (par capillarité). ? Elles sont l’explication de la qualité de réservoirs d’eau des sphaignes.
Les feuilles vont avoir un développement le long de la tige ou perpendiculaire à la tige.
2 Les Bryales.
Les bryales sont divisées en deux familles : les bryales acrocarpes et les bryales pleurocarpes.
Chez les acrocarpes, les archégones sont en position terminale. Chez les pleurocarpes, les archégones sont latéraux, sur la tige. Ces dernières ont aussi, généralement, des tiges couchées, contrairement aux acrocarpes qui ont des tiges dressées.
La famille des polytriches ou des Polytrichaceae. Genre Polytrichum.
Les plantes de ce genre sont souvent de bons indicateurs des conditions d’un milieu. Par exemple :
- Polytrichum commune vit dans les milieux humides, voir tourbeux. Elle est d’assez grande taille.
- Polytrichum juniperynum. Cette plante est typique des sols secs, voir très secs.
- Polytrichum formosum est une espèce des milieux humides, qui se dessèche rapidement.
V Les ptéridophytes : classification, écologie, utilisation.
Les Psilophytinées sont des cas très primitifs. Il n’en reste qu’un seul genre (Psilotum).
Les Lycopodinées. Elles ont une tige aérienne rampante et un aspect mousseux. Elles existent aussi sous forme aquatique.
A Les Lycopodinées.
Elles ont des sporanges regroupés à la base des feuilles.
La famille des Isoètes est généralement immergée. Elles ont des allures de graminées. A la base des frondes, on a un renflement qui correspond au sporange. Ce denier donne un microsporange ou un macrosporange. Dans les deux cas, le gamétophyte reste dans les feuilles.
Chez certaines espèces d’Isoètes, il y a peu de stomates et la cuticule est très épaisse au niveau des feuilles. Le CO2 pour la photosynthèse est pris dans les sédiments.
Tous les lycopodinées ont un aspect de mousse.
1 Les lycopodiaceae.
Tous les sporanges sont semblables. Le genre Lycopodium est dominant. Les sporanges sont regroupés à une extrémité et forment des épis.
Ces plantes sont beaucoup utilisées en pharmacie ; pour les poudres de lycopodes, faites à partir des spores. Elles ont des propriétés adoucissantes et anti-inflammatoires. Elles sont aussi utilisées pour faire des pilules.
Au XVIIème et au XVIIIème siècle, les lycopodes étaient employés pour les feux d’artifices.
2 Les Sélaginellaceae.
Les sporanges sont à l’aisselle des feuilles. Il va s’en différencier des macrosporanges et des microsporanges.
B Les Equisetinées.
La tige souterraine porte des tiges aériennes, articulées, dressées, avec des feuilles verticillées.
Les Equisetaceae sont un ordre des Equisetinées. On trouve par exemple Equisetum arvena, la prêle. Les feuilles sont extrêmement réduites et forment une gaine autour de la tige (articulée).
On trouve des tiges fertiles (vernales et printanières) qui ont des sortes d’épis à l’extrémité, contenant des sporanges. Les autres tiges sont stériles. Les deux types de tiges sont riches en silice.
C Les Filicinées.
Les filicinées sont les fougères actuelles. Elles n’ont pas de feuilles mais des frondes.
A l’intérieur, on peut distinguer les Ophioglossaceae, à fronde solitaire ; les Polypodiaceae à frondes plus nombreuses ; les Osmundaceae et les Hydroptérydales, plantes aquatiques ou semi-aquatiques.
1 Les Ophioglossaceae.
Deux cas sont possibles.
- Soit, une fronde entière : c’est le genre Ophioglossum, relativement rare. Les plantes de ce genre mesurent 10 à 20 centimètres de haut et poussent dans les prairies humides. Elles sont rares car les prairies humides sont rares.
- Soit, on trouve une fronde stérile découpée et une fronde fertile avec des sporanges. C’est le genre Botrychum. Ces plantes mesurent 10 centimètre de haut et sont en montagnes (exemple : Botrychum lunaria). Botrychum loceolatum est une espèce disparue de France. En 1926, elle était observée dans toute la région du Mont Blanc. En 1943, il n’en restait qu’un seul pied. Elle a disparu à cause des botanistes. Botrychum multifidum est la plante la plus rare de France. Elle n’existe qu’en un endroit et présente un ou deux pieds tous les deux ans.
2 Les Polypodiaceae.
Parmi les Polypodiaceae, on trouve la fougère aigle (Ptéridium aquilinum) qui existe sur les 5 continents. Ce sont des plantes qui repartent après les feux.
3 Les Osmundaceae.
Osmunda regalis est la plus grande fougère de France : elle mesure entre deux mètre et deux mètre cinquante. Elle pousse en milieu humide, au bord des ruisseaux.
4 Les Hydroptérydales.
Ce sont des fougères aquatiques.
La famille des Marsileacées, genre Marsilea. Les espèces de ce genre ressemble à des trèfles à quatre feuilles mais n’en sont pas.
Il y a existence de sporocarpes au niveau du rhizome qui permettent la reproduction de ces fougères.
Exemple : Marsilea quadrifolia strigosa, est une espèce que l’on ne trouve que dans l’Hérault, avec seulement trois populations.
VI Les gymnospermes.
Les gymnospermes font parti de l’embranchement des spermaphytes. Ils n’ont pas de fruit.
Ce sont des végétaux ligneux, souvent arborescents. Toutefois, il existe des genévriers rampants, de 10 centimètres de haut.
Le limbe des feuilles est généralement peu développé : petites feuilles en aiguilles ou en écailles.
Les gymnospermes sont très minoritaires par rapport aux angiospermes. Toutefois, ce sont des arbres de ce sous-embranchement qui possèdent divers records :
- Record de hauteur : Séquoia, 120 mètres.
- Record de taille : Taxodium, 6 mètre de diamètre.
- Record de longévité : Pin de 4600 ans aux USA, Pinus longaeva.
Ce sous-embranchement est divisé en :
- Cycadophytes : pré-spermaphytes, avec Ginkgo.
- Coniférophytes : les vrais gymnospermes sensus stricto.
- Gnétophytes : ce sont des clamidospermes, des intermédiaires entre les gymnospermes et les angiospermes.
A Les Cycadophytes.
Chez les cycadophytes, il y a encore zoïdogamie : la fécondation peut encore avoir lieu alors que l’ovule présente déjà des réserves.
On se trouve chez des espèces dioïques (pied mâle et pied femelle).
La « fleur » male constitue un cône, constitué de centaines de microsporophylles. A la surface inférieure de ces microsporophylles, on trouve des microsporanges qui libèrent des grains de pollen.
Le « cône » femelle est constitué de mégasporophylles qui portent à leur base, des ovules. Le cône femelle est l’équivalent de la fleur femelle des angiospermes.
Au moment de la fécondation, les grains de pollen nagent dans le liquide sécrété par l’ovule. Après cette fécondation, il y a développement embryonnaire, formation de la plantule ? On n’a pas de véritable graine car on n’assiste pas à une quelconque dormance.
Les Cycadopsides.
Ordre : Cycadales ; Famille ; Cycadacées.
• Le genre des Cycas.
L’allure des Cycas est proche de celle du palmier. Ce sont des plantes de petite taille ; le tronc est court et les feuilles sont pennées.
Les Bennettitales.
Les bennettitales sont un groupe de fossiles, éteints au crétacé.
B Les Ginkopsides.
Les ginkopsidés sont des arbres dioïques. Les étamines sont portées par un pédoncule. La fleur femelle est extrêmement réduite. Les plantes de ce groupe ont conservé une certaine dichotomie.
C Les Coniféropsides.
1 F : Pinales / O : Pinacées.
Ces végétaux ont des feuilles en aiguilles alternes. Elles sont en aiguilles ou linéaires, généralement persistantes (exception pour le mélèze).
Les cônes femelles sont ligneux. Les macrosporophylles portent deux ovules. Une écaille porte deux ovules, avec une bractée indépendante de l’écaille.
Cette bractée est importante car elle montre que cet ensemble correspond à une fleur. Le cône femelle de pin est une inflorescence.
Organisation de l’appareil végétatif.
Un Abiès (sapin) montre une organisation différente de celle d’un cèdre ou d’un pin : on a trois types différents. Les différences portent sur les rameaux et les feuilles.
L’organisation la plus originale est celle des pins. Les rameaux courts (brachiblastes) portent les aiguilles (les pseudophylles) qui sont des rameaux transformés. Les vraies feuilles sont sur les brachiblastes (en écailles) mais ne sont pas chlorophylliennes.
Le genre Pinus.
• Pinus sylvestris.
Ce pin a une écorce typique. La partie supérieure du tronc est orangée. Les pseudophylles sont de petite taille. Les cônes sont très petits.
C’est une espèce majeure car elle est l’espèce principale de la taïga (forêts boréales de hautes altitudes). Cela représente le tiers de la surface forestière mondiale.
Un peu plus haut vers le nord, il n’y a pas d’arbre, c’est la toundra.
• Pinus uncinata (Le pin à crochets).
Ce pin porte de petites aiguilles et de petits cônes. Le tronc n’est jamais orangé. Au niveau des cônes, l’écusson possède un crochet (écusson recourbé en crochet).
Pinus uncinata est parfois introgressé avec Pinus sylvestris ? zones d’hybridation.
Pinus uncinata est uniquement présent en haute montagne. ? Il constitue la limite supérieure de la forêt (entre 2,5 et 3 kilomètres).
• Pinus cembra (Le pin cembro ou arolle).
Ce pin est caractéristique des alpes. Il est reconnaissable car il présente des pseudophylles aux brachiblastes. Il monte jusqu’à 2,5km d’altitude. (P. cembra = P. siberica).
Les graines de ce pin sont relativement lourdes et ont donc un problème de dissémination. Les oiseaux de type Nucifraga servent à leur dissémination (80 à 90% des graines). Ils tapent sur les pédoncules et détachent les cônes de l’arbre qu’ils vont emporter à une certaine distance, sur une « enclume » naturelle. Ils décortiquent les cônes. Ils avalent les graines ou les stockent en les enfouissant par paquet, dans l’humus du sol. Il reste toujours des stocks oubliés qui permettront la germination de nouveaux individus.
• Pinus longaeva (USA).
Ces pins se développent entre 3000 et 3500 mètres d’altitude, dans les environnements arides. Le plus vieux des P. longaeva est âgé de 4767 ans. Le plus grand d’entre eux fait 11 mètres de circonférence.
Les aiguilles peuvent vivre une trentaine d’années, ce qui est une considérable économie d’énergie : métabolisme restreint. On observe que peu de xylème (bois) est fonctionnel : les besoins énergétiques sont donc moins importants. Ces arbres ne connaissent pas les invasions de bactéries, d’insectes, de champignons grâce à la grande quantité de résine. Ils sont très espacés les uns des autres et ceci leur permet d’avoir un système racinaire très développé.
Les fleurs et les fruits ne sont formés que lorsque l’arbre est relativement âgé.
• Pinus pinaster : le pin maritime.
Ce pin est de couleur vert foncé, piquant et extrêmement gros. Il existe à l’état spontané et a beaucoup été planté dans la forêt landaise. Cette forêt fait environ un million d’hectares (100km ? 100km). C’est une forêt artificielle (à l’origine c’était une chênaie). Elle fut plantée au 19ème siècle. En 1945 et 1949 elle subit des incendies.
• Pinus halopensis (le pin d’Alep).
Ce pin est méditerranéen, adapté à ce climat. Il est reconnaissable à ses aiguilles et son feuillage léger. Les cônes sont groupés par deux ou trois. Les pins sont des espèces héliophiles.
• Pinus coultheri.
Ce pin présente des cônes très longs, entre 30 et 40 centimètres de long.
Le genre Abies (sapin).
• En France, on a une espèce spontanée : Abies alba aussi appelée Abies pectinata. C’est une espèce typique de l’étage montagnard (sapin et hêtre). Les sapins sont fréquemment utilisés pour faire des bateaux, d’où les diverses sapinières.
Remarque : En Andalousie, Abies pinsape pousse sur la sierra.
• Picea abies (Epicéa).
Cette espèce n’est pas spontanée dans les Pyrénées mais il en a existé il y a 20 000 à 30 000 ans. Les pessières sont des plantations d’épicéa (qui existent dans la montagne noire). L’épicéa est classique dans les alpes et les vosges.
Les différences entre l’épicéa et les sapins :
- Les sapins ont des branches beaucoup plus horizontales.
- Les cônes restant, s’ils sont entiers et allongés, proviennent sûrement d’un épicéa car les cônes de sapins se désagrègent sur l’arbre et il n’en reste que des écailles. Le cône est dressé les sapins chez, pendant chez l’épicéa.
- Le feuillage montre deux ramifications latérales chez les sapins et six chez l’épicéa.
- Les rameaux ont un aspect rugueux chez l’épicéa car il reste les coussinets qui portaient les aiguilles.
L’épicéa est comme le pin sylvestre, typique de la forêt boréale (taïga) : les aiguilles d’épicéa restent vivantes en dessous de –30°C et peuvent assimiler jusqu’à –10°C.
Dans les alpes, l’épicéa permet de donner des populations clonales par marcottage (multiplication végétative).
Le genre Cedrus.
• Cedrus deodora (Himalaya) ; Cedrus atlantica (Maroc, Algérie) ; Cedrus libani (Liban).
Les caractères de différenciation des cèdres sont la longueur des aiguilles et la grosseur du cône. Le cèdre du Liban n’existe plus que sur 70 à 80 hectares et a des problèmes de régénération.
2 F : Pinales, O : Cupressacées.
Le genre Juniperus.
Les cupressacées sont à feuilles opposées ou verticillées, en écailles ou en aiguilles/cône bacciforme ou arrondi-cubique.
On trouve des cônes en forme de baies. Les cônes charnus (les galbules) sont caractéristiques du genre Juniperus. Les écailles deviennent charnues.
Le passage de ces galbules par le tube digestif d’animaux est obligatoire pour la dissémination.
• Juniperus communis.
Il possède des feuilles en aiguilles piquantes. On le trouve un peu partout. Ses galbules sont bleutées à maturité. La feuille montre une bande de stomates.
• Juniperus oxycedrus.
Il possède les mêmes feuilles piquantes que le genévrier commun. Une différence tout de même, on trouve deux bandes de stomates et non une. Les galbules sont rouges. Ce sont des plantes méditerranéennes. Elles servaient à faire de l’huile de cade.
• Juniperus thurifera.
C’est un arbre aux dimensions impressionnantes. Il participe à la constitution de la limite supérieure de la forêt (? 2800 mètres d’altitude). Ces Juniperus thurifera peuvent atteindre plus de 2 mètres de diamètre. Ils n’ont pas de feuilles en aiguilles mais en écailles, identiques aux feuilles de cyprès. Les feuilles ne sont jamais alternes. Le cône est charnu.
Le genre Cupressus.
Ce sont les cyprès. On trouve Cupressus sempervirens (dans les cimetières) ; Cupressus dupreziana qui n’existent que dans le sud de l’Algérie, il n’en reste que quelques centaines d’individus.
3 F : Pinales ; O : Taxodiacées.
Exemple : Taxodium distichum. C’est le cyprès chauve : il perd ses feuilles en hiver. On note la présence de pneumatophores (racines aériennes).
D Les Taxopsides.
Le genre Taxus.
Ce genre est parfois classé avec les conifères, pourtant, il n’a pas de canaux sécréteurs de résine.
L’appareil végétatif est comparable à celui d’Abies. Par contre, la graine est complètement différente : l’arille, enveloppe charnue rouge, porte la graine.
C’est une espèce très toxique (des feuilles et des graines).
Le taxol en est extrait ; c’est une substance anticancéreuse. Elle est présente chez Taxus breviflora, dans l’écorce.
Chez Taxus baccata, le taxotère peut être isolé du feuillage et il est encore plus puissant que le taxol.
E Les Gnétopsides.
Ces gnétopsides sont parfois complètement exclus des gymnospermes et forment alors le groupe des Clamidospermes.
La graine commence à être entourée par une sorte d’enveloppe.
On y trouve trois familles importantes.
1 Les Gnétacées ; genre Gnetum.
On trouve ces plantes dans les forêts tropicales humides. Leurs feuilles ressemblent à celles d’angiospermes.
2 Les Ephedracées ; genre Ephédra.
On les trouve sur l’arrière des dunes des plages. Leurs rameaux sont articulés. Elles peuvent être des espèces rupicoles (poussant sur les rochers).
3 Les Welwitschiacées ; genre Welwitschia.
Il n’existe ici qu’un genre avec une unique espèce : Welwitschia mirabilis.
Elle n’existe que dans le désert du Kalahari (au sud de l’Afrique) : milieu extrêmement sec. L’appareil végétatif ne présente que feuilles dilacérées. Elle peut aller chercher l’eau jusqu’à 75 centimètres de profondeur. Grâce à ses deux feuilles, très larges, elle peut capter l’eau des rosées.
VII Les angiospermes.
A La classification des Angiospermes.
Les analyses moléculaires ont montré que l’ancienne classification mettait ensemble des plantes sans grand rapport génétique.
On différencie maintenant les angiospermes en Eudicotylédones et Monocotylédones. Certains dicotylédones se sont retrouvés avec les monocotylédones.
B La classe des monocotylédones.
On trouve sept super-ordres.
a Morphologie foliaire.
Dans la grande majorité, les feuilles sont simples, à nervures parallèles. Dans certains cas, on trouve des feuilles proches de celles des dicotylédones.
Les dioscoréacées et le palmier (palmacées) ont des feuilles composées complexes.
b Caractère embryonnaire et plantule.
Les monocotylédones n’ont qu’un cotylédon.
c Anatomie.
Les vaisseaux vasculaires sont nombreux et répartis uniformément dans le cylindre central (= atactostèle).
Il n’y a jamais de cambium, donc pas de formations secondaires (enfin, presque jamais).
Remarque : certains dicotylédones sont aussi atactostèles (les Pipérales).
d Morphologie florale.
Les fleurs sont généralement trimères. L’ovaire est supère, altère.
Ce type de fleurs trimères existe aussi chez les magnoliales, dicotylédones primitifs, ne faisant pas parti des eudicotylédones.
e Les apertures polliniques.
L’aperture pollinique est un amincissement de la paroi qui permettra la pollinisation. Chez les monocotylédones, les sacs polliniques sont mono-aperturés.
1 Le super-ordre des Liliiflorae.
Ce super-ordre regroupe beaucoup de plantes ornementales et alimentaires. Avant on distinguait deux ordres : les Liliales (Liliaceae) et les Orchidales (Orchidiaceae).
Les Liliales ont des fleurs actinomorphes. Les étamines sont normales, non soudées au gynécée. La graine est grosse, avec beaucoup d’albumen.
Les Orchidales ont des fleurs zygomorphes. Les étamines sont gymnostèmes (soudées) et la graine est minuscule, sans albumen.
Les Liliales sont divisées en Iridaceae ou Amaryllidaceae (ovaire inférieur) et en Liliaceae (six étamines et ovaire supère).
a L’ordre des Asparagales.
Exemple de la famille des Agavaceae.
Cette famille contient des yuccas et des agaves. Avant, le yucca faisait parti des Liliacées et les agaves des amaryllidaceae. Ce classement était fait car les yucca ont un ovaire supère et les agaves l’ont infère. Maintenant, ils sont regroupés car
- L’appareil végétatif est composé de feuilles un peu grasses, épineuses, charnues.
- L’écologie est assez proche : milieu aride.
- Ils ont tous deux des formations secondaires.
- Leur garniture chromosomique est originale : N=30.
La position de l’ovaire semble, dans ce cas, un mauvais marqueur phylogénique.
Les agaves ont une unique floraison. Après celle-ci, c’est la mort de la plante.
Le mescal est une boisson obtenue à partir de la sève de certains agaves.
b L’ordre des Dioscoreales.
Cet ordre est original dans les monocotylédones : les feuilles sont différentes des autres monocotylédones ? elles sont cordiformes.
Les dioscoreales sont presque toujours des lianes (plantes volubiles). Les espèces sont dioïques.
Remarque : Sur 13500 genres de plantes à fleur, 7% des genres contiennent des espèces dioïques. Sur 240 000 espèces, 6% sont dioïques. Parmi les espèces dioïques, beaucoup sont des lianes.
Les dioscoréales contiennent trois genres principaux.
1 Le genre Tamus.
Exemple : Tamus communis, « l’herbe aux femmes battues ». Cette plante a des propriétés anti-contusions venant du rhizome. Elle provoque des traces rouges si elle est appliquée sur la peau.
2 Le genre Diascorea.
Les fleurs mâles sont composées de 6 tépales, 6 étamines. Les fleurs femelles portent 6 tépales, six étamines vestigiales et un ovaire divisé en trois loges.
On a une trimérie : 3+3 tépales.
3 Le genre Borderea.
Le genre Borderea est plutôt tropical. C’est une plante rupicole.
Les espèces de Borderea sont des survivances du tertiaire (climat tropical).
Elles sont herbacées, perdent leurs feuilles et laissent des écailles sur le haut du bulbe.
On trouve deux exceptions : Borderea pyrenaica et Borderea chouardii qui n’existent que dans les Pyrénées.
Les dioscoreales comprennent environ 60 espèces.
L’Igname est utilisée pour la consommation (son tubercule) et pour la synthèse de DHEA (enzymes, protéines).
c L’ordre des Liliales.
Les liliales sont des herbes vivaces, possédant des organes souterrains de réserves (rhizomes, bulbes ou tubercules).
Au niveau de la fleur, par exemple chez les muguets, on a 6 tépales (3+3 tépales pétaloïdes), 6 étamines et 3 carpelles à ovaire supère. La fleur est régulière : actinomorphe.
L’espèce Fritillaria meleagris présente une corolle en damier.
c1 La famille des Liliaceae.
1 Le genre Tulipa.
Toutes ces plantes sont des espèces originaires du proche orient. Tulipa gesneriana est la tulipe cultivée classique. On trouve quelques tulipes « sauvages » qui deviennent de plus en plus rares. Les tulipes cultivées ont été introduites au 16ème siècle.
Les formes sauvages sont différentes : elles sont soit indigènes, soit furent introduites plus tôt que l’espèce cultivée (invasion sarrasine, croisades…)
Les trois espèces suivantes sont des espèces menacées :
- Tulipa platystigma : il n’en reste que quelques pieds dans les Hautes Alpes.
- Tulipa agenensis : ce sont des adventices des cultures, c’est à dire, des plantes qui se développent en association avec les cultures (dans les champs cultivés). Cela correspondait à des pratiques culturales traditionnelles maintenant abandonnées. ? forte régression.
- Tulipa praecox : c’est aussi une espèce adventice. Elles furent combattues car elles étaient trop envahissantes. Maintenant, elles sont prêtes à disparaître.
Ce sont des plantes liées à l’Homme.
2 Le genre Lilium.
Le lys.
c2 La famille des Iridaceae (Iris, glaïeuls, crocus).
Les Iridaceae sont généralement des herbes vivaces, avec des rhizomes, des bulbes ou des tubercules. La fleur peut être actinomorphe ou zygomorphe. L’ovaire est infère. On trouve 3 étamines et 3+3 tépales. La fleur d’Iris a des stigmates pétaloïdes.
Exemple : Crocus sativus (le safran).
Le condiment correspond aux stigmates. La culture de cette plante est longue (une livre de safran = 35 000FF). Crocus sativus est originaire d’Asie mineure. Le safran était cultivé 500 à 600 avant JC. Il fut introduit en Europe par l’intermédiaire des conquêtes arabes. La forme sexuée ne se reproduit que par multiplication végétative ? La fleur est stérile.
Au moyen-âge, la fraude au safran valait la pendaison. Cette fraude se fait avec du carthame (Cartamus) car la fleur ressemble aux stigmates de safran.
c3 La famille des Orchidaceae.
Les orchidaceae sont des herbes terrestres ou épiphytes (vivant dans les arbres). Elles peuvent être saprophytes.
La fleur est zygomorphe. On trouve 3+3 pétales dont un qui forme l’éperon. L’éperon va avoir un rôle dans la pollinisation. Le filet des étamines, le style, le stigmate sont soudés et constituent le gymnostème.
Les grains de pollen sont groupés au niveau de pollinies (deux).
Les graines sont minuscules, sans albumen ? La double fécondation ne fonctionne pas.
Il va y avoir mycotrophie : symbiose avec un champignon qui permet le développement de l’embryon pendant la germination. Le champignon concerné est du genre Rhizoctonia.
La graine est constituée d’environ 100 à 150 cellules indifférenciées, infestées par les filaments du champignon.
La mycorhize est nécessaire pour la croissance de la plante et aide le champignon par les apports de substances carbonées.
Les pièces florales sont adaptées à la pollinisation entomophile (par les insectes).
On trouve deux groupes principaux : Orchis et Ophrys (un pétale ressemble à un insecte).
D’un point de vue alimentaire, chez la vanille, la « gousse » vient du développement de l’ovaire infère.
2 Le super-Ordre des Commeliniflorae.
a L’ordre des Poales.
La famille des Graminées (gramineae).
Les graminées ont une tige particulière (chaume) creuse sauf au niveau des nœuds. Elle est cylindrique.
Les feuilles sont distiques (insérées sur deux rangs). L’inflorescence de base est un épillet. L’épillet est un ensemble de feuilles réduites, situées au dessus de deux pièces (les glumes). Le fruit est particulier : c’est un caryopse, un fruit sec indéhiscent où le péricarde adhère à la graine.
Un certain nombre de formations comme les steppes, les savanes, les prairies, les pelouses et les pampas sont dominées par les graminées. Ce sont souvent des formations de dégradation.
Par exemple, dans le bassin méditerranéen, on a des steppes à Alfa (Stipa stenacissima) qui couvrent des millions de km². Cet Alfa sert à la fabrique de pâte à papier, de tapis… Dans les steppes, on a des touffes de graminées séparées (discontinues). Il y aurait des arbres dans ces zones (Afrique du nord) de plus les steppes sont sur-exploitées.
Les dunes littorales. Ce sont des dunes mouvantes avec une graminée bien adaptée : Ammophila arenaria (l’oyat). Ce sont des milieux peu dépendant du climat général. L’oyat va former un système de rhizomes très performant (réseau dans la dune) qui filtre et ralentie les mouvements des dunes.
Les pelouses calcaires sont des formations de dégradation, dominées par les graminées.
Les graines de graminées constituent l’essentiel des céréales. Le sarrasin (ou blé noir) n’est pas une graminée mais un Polygonacées (Fagopyrum), un dicotylédone.
La cane à sucre est Saccharus officinarum ; Zea mais est le maïs.
a1 Le genre Triticum (les blés).
On trouve trois groupes différents.
• Les engrains, Triticum monococcum.
Ce sont les blés les plus anciens. Ils ont 2N = 14 chromosomes. Ils sont diploïdes.
• Les blés durs, Triticum durum.
Ils ont 2N = 28 chromosomes. Ils sont largement cultivés ; servent aux pâtes alimentaires.
• Les blés tendres, Triticum aestivum ou T. vulgare.
Ils ont 2N = 42 chromosomes. Ils sont utilisés pour la fabrication du pain.
Trois graminées sauvages seraient à l’origine de ce blé actuel, dont deux du genre Aegylops.
a2 Le genre Oriza, le riz : Oriza sativa.
Le riz est l’espèce la plus cultivée après le blé. Environ 6000 variétés de riz sont connues. Oryen glaberrima est une espèce d’Afrique occidentale.
Le riz récolté est le riz paddy : c’est un caryopse (fruit) avec les glumes et les glumelles. Quand il n’y a que le caryopse, on parle de riz brun ; Le riz brun auquel on a enlevé le péricarpe est le riz blanc. Le péricarpe contient des vitamines A et B.
b L’ordre des Cypérales.
La famille des cypéraceae.
Les épis de cypéraceae sont généralement unisexués. Le genre principal est Carex (la laîche). On trouve aussi le genre Cyperus qui comprend des plantes de milieux humides (Cyperus papyrus).
3 Le super-ordre des Areciflorae
L’ordre des Arecales
La famille des Palmae = Arecaceae.
C’est chez les Palmae que l’on trouve les palmiers. On trouve environ 2500 espèces.
Ils ont la taille d’arbres. Le tronc est formé par la soudure de la base des feuilles. Les feuilles sont dites palmées ou pennées.
Elæis guineensis est le palmier à huile.
Phœnix dactylifera est le palmier dattier (les dattes sont sur les pieds femelles). Dans les palmeraies cultivées, on trouve 90% de pieds femelles pour 10% de pieds mâles. Dans la nature, on trouve un équilibre de 50% de chaque sexe.
Le palmier nain (Chamærops humilis) sert pour fabriquer de la ficelle… Il est représentatif du thermo-méditerranéen : en dessous de Barcelone.
Le cocotier (Coccos nucifera) donne la noix de coco qui est une drupe dont la dissémination se fait par hydrochorie. Il sert aussi à donner le coprah.
4 Conclusion sur les Monocotylédones.
Les monocotylédones ont des feuilles à nervures parallèles. La fleur est trimère.
Ils constituent une réussite extraordinaire au niveau morphologique, écologique (en biodiversité)… Par exemple, les Orchidaceae regroupent 750 genres différents et 18000 espèces distinctes.
Actuellement, les gymnospermes comprennent 800 espèces. A elles seules, les graminées présentent 660 genres pour 8000 à 10 000 espèces. Les cypéraceae comprennent 90 genres pour 4000 espèces.
Les monocotylédones sont un groupe très diversifié, avec de fortes divergences :
- Les Orchidaceae présentent une fleur élaborée (pour la zoogamie), des graines minuscules et innombrables, des graines fragiles.
- Les Poaceae montrent des fleurs petites, discrètes avec une grande quantité de pollen (anémogamie). Les graines sont albuminées, souvent solitaires. L’inflorescence est particulière (épis et épillets). Les graines sont résistantes.
C Les quatre ordres de dicotylédones archaïques.
Ces dicotylédones archaïques (ou paléoarbres/paléoherbes) sont regroupés avec les monocotylédones. Ils sont dits archaïques car ils ont un pollen uni-aperturé, des vaisseaux imparfaits, des fleurs encore trimères et une disposition spiralée des pièces florales.
1 Super-ordre des Magnoliales, Ordre des Magnoliaceae.
Magnolia/Liriodendron tulipifera.
Ce sont des espèces, introduites en France, qui possèdent des caractères archaïques :
- Disposition spiralée des pièces florales.
- Anthères à filet large qui donne des étamines lamellaires.
- Le nombre des étamines est non fixé.
- Le carpelle donne un follicule (fruit déhiscent) qui continue sur une graine suspendue au funicule.
- Il n’y a pas de différenciation entre mes sépales et les pétales : que des tépales. Le périanthe est homochlamydé.
- On ne trouve que des trachéides : le bois est homoxylé, comme les gymnospermes. (les espèces à bois hétéroxylé ont des vaisseaux parfaits et des fibres).
2 Les Iliciaceae.
On trouve Illicium anisatum : la badine ou anis étoilée.
3 Les Laurales/Ordre des Lauraceae.
C’est une famille d’arbres ou d’arbustes odorants des régions tropicales ou sub-tropicales. Les feuilles sont simples. On trouve entre autre Laurus nobilis, le laurier sauce.
Remarque sur les autres "Lauriers" :
- Nerium oleander, le laurier rose fait partie des Apocynaceae ;
- Virburnum tinus, le laurier tin fait partie des Caprifoliaceae ;
- Prunus laurocerasus, le laurier cerise, fait partie des Rosaceae.
Ce sont des plantes triplostémones : trois fois plus d’étamines que le nombre de base. Les étamines ont un clapet. La feuille est tétramère (Laurus nobilis). Persea americana est l’avocatier ; donne les avocats qui sont des baies. Cinnamonum verum est le cannelier : Cinnamonum camphora est le camphrier.
D Les Eudicotylédones.
Les eudicotylédones sont aussi appelés angiospermes supérieurs.
On trouve parmi eux, des cas à caractères archaïques, comme : la spiralisation, la présence de tépales, des cas de trimérie et la dialycarpie.
L’évolution va dans le sens de la soudure des pièces.
1 Les Ranunculales.
a Les Papaveraceae.
Les papaveraceae ont toujours du latex. Les fleurs sont tétramères, gamocarpellées.
Elles sont toujours herbacées et présentent deux sépales qui deviennent rapidement caduques.
Les quatre pétales ont une préfloraison chiffonnée.
L’ovaire est supère, uniloculaire à placentation pariétale.
Le fruit est particulier : c’est un pyxide. Il s’ouvre par des pores apicaux. La chélidoine (Chelodonium majus) a un fruit déhiscent, de type silique avec diminution du nombre de carpelles.
Coquelicot = Papaver rheas ; Pavot = Papaver somniferum.
L’opium est obtenu à partir d’une incision dans la capsule du pavot (trois semaines après la fécondation) : un latex va s’écouler et coaguler. On peut en faire de la morphine ou de la codéine. Si l’on rajoute deux groupements acétyles, on obtient de l’héroïne. Les graines de pavot sont consommables.
b Les Ranunculaceae.
Les ranunculaceae n’ont pas de latex. Les fleurs seront trimères ou pentamères et généralement dialycarpellées.
Ce sont des herbes pérennes (vivaces) ou annuelles. Les fleurs sont spiralo-cycliques : le périanthe a des pièces spiralées et/ou cycliques. Les fleurs sont actinomorphes ou zygomorphes (Aconitum est zygomorphe). Les ranunculaceae sont toujours dialytépales ou dialypétales/dialysépales.
On trouve de nombreuses étamines (polystémonie) en nombre non fixe.
Les carpelles sont plus ou moins nombreux ; l’ovaire est supère.
Les fruits sont des akènes ou des follicules.
Les ranunculaceae sont peu évoluées mais présente une tendance à la gamocarpellie (cas de Nigella : trois carpelles soudés) et à la diminution du nombre de carpelles.
Les plantes zygomorphes commencent à exister (exemple : Aconitum).
C’est une famille avec des plantes toxiques voir mortelles ; beaucoup de Ranunculus sont toxiques. Aconitum vulparia (aconit tue-loup) contient de l’aconitine (alcaloïde) dans les racines. Elle est le cas le plus toxique en Europe. Toutes les parties de la plante sont toxiques. L’ingestion de 2 à 4 grammes de racines est mortelle.
L’Hellébore a des graines toxiques.
Le développement de nombreuses renoncules se fait en milieu humide.
2 Les Caryophyllales.
Dans la graine, l’embryon est courbe, voir même enroulé, en fer à cheval. Il s’enroule autour des tissus de réserve ? centrospermales (= Caryophyllales).
De nombreuses espèces sont des xérophytes.
On trouve aussi des plantes halophytes, comme la famille des chénopodiaceae.
a Les Cactaceae.
Les cactaceae sont une famille presque exclusivement américaine. En Europe, elles ont été introduites par l’Homme.
Cactus n’existe pas comme genre.
Ce sont toujours des plantes succulentes dont les tissus font de la rétention d’eau. Elles sont généralement épineuses. Elles peuvent être en forme de cierge, de raquette ou en « coussin de belle-mère ».
Seul le genre Pereskia possède des feuilles. Habituellement, elles sont transformées en épines.
Les tiges présentent des aréoles (petites dépressions) souvent remplies de poils, avec la sortie des épines.
Les fleurs sont à nombre indéfini de pièces : nT + nE + 3 à nC.
Opuntia ficus indica (figuier de barbarie) présente des raquettes charnues. Il joue un grand rôle dans les régions d’Afrique du nord. Sa fleur a la même formule que précédemment. L’ovaire est infère et, une fois fécondé, il donne un faux fruit. C’est un ovaire uniloculaire à placentation pariétale.
Dans cette famille, on trouve Peyotl : Lophophoria échinocactus qui contient de la mescaline : hallucinogène puissant. Le peyotl n’est pas épineux et pousse dans le nord de l’Amérique.
b Les Caryophyllaceae.
Ce sont des plantes herbacées à feuilles opposées, portées au niveau des nœuds, renflés. L’inflorescence est une cyme bipare. La fleur est pentamère avec dix étamines (double du nombre de base ? diplostémonie). Le cycle externe d’étamines est en face des pétales ? obdiplostémonie.
Les carpelles sont généralement au nombre de cinq, à placentation centrale : ovaire uniloculaire.
Dans cette famille, on distingue deux tribus (sous-familles) :
- Tribu à sépales libres : les Alsinées. Exemples : Stellaria, Cerastium.
- Tribu à sépales soudés : les Silénées. Les sépales se soudent en un tube. Le limbe sort de ce tube par un onglet. Exemples : Dianthus (œillet), Lychnis, Silène, Saponaria.
Ce sont des plantes liées aux moissons (messicoles), comme par exemple, la nielle (Agrostemma githago).
3 Les Fabales.
Les Fabaceae (= Légumineuses).
Les fabaceae sont une famille homogène car un caractère est typique : le fruit, ou le « légume », est une gousse. La gousse est un fruit dérivant d’un seul carpelle et s’ouvrant habituellement par deux valves (une ventrale et une dorsale).
Ces légumineuses abritent des bactéries, fixant l’azote atmosphérique, au niveau de racines : les nodosités (bactéries du genre Rhizobium).
Ce sont des plantes pouvant coloniser de nombreux milieux grâce à ces nodosités. Les feuilles sont composées.
a Première sous-famille.
La fleur est actinomorphe. Les fleurs sont petites, groupées en inflorescence compacte avec n étamines. ? Mimosoïdae. Ce sont toujours des arbres ou des arbustes.
L’appareil végétatif est constitué de feuilles composées de nombreuses folioles. Dans certains cas, les feuilles arrivent à disparaître et ce sont les phylums – rachis aplatis – qui assurent la fonction chlorophyllienne. Chez certaines familles, on trouve souvent des stipules, petits appendices à la base des feuilles. Ces stipules peuvent être transformées en épines.
La fleur : Chaque petite boule correspond à une inflorescence. C’est une fleur actinomorphe régulière avec de nombreuses étamines.
Le genre Acacia.
Les Acacias sont africains et australiens. En France, ce que l’on appelle mimosa est du genre Acacia. On trouve des espèces africaines spontanées, typiques des milieux sub-désertiques. Quand il fait trop chaud et trop sec, les arbres perdent leurs feuilles.
Acacia senegal donne la gomme arabique.
Mimosa pudica a des folioles qui se referment quand on les touche.
On trouve aussi les albizzias.
La symbiose Acacia/fourmis ou myrmécophilie.
Certains Acacias hébergent, dans leurs stipules, des fourmis. Acacia conigera abrite des fourmis Pseudomyrmex ferrugina. Ces fourmis sont nourries par l’acacia. En retour, elles débarrassent l’arbre des plantes épiphytes et le protègent contre les herbivores.
Des Acacias proches de conigera existent mais sont sans fourmis ; ils ont des défenses chimiques (cyanogènes) à la place.
bSeconde et troisième sous-familles.
Dans les deux cas, les fleurs sont zygomorphes avec jusqu’à dix étamines.
• Les Faboïdeae.
La préfloraison est vexillaire ; présence de dix étamines (diadelphes).
Les feuilles sont composées, pennées. On note la présence de stipules parfois modifiées en feuille ou vrille.
La fleur est zygomorphe avec une corolle papillonacée, typique de la préfloraison vexillaire. Cette fleur est pentamère avec : un pétale supérieur formant l’étendard, deux pétales latéraux (les ailes) et deux pétales inférieurs (la carène).
Les étamines sont au nombre de dix (diplostémonie). Elles sont soit toutes soudées entre par le filet et forment le tube : androcée monadelphe, soit neuf étamines sont soudées et une est libre (à côté de l’étendard) : androcée diadelphe.
On trouve un carpelle à ovaire supère qui donne une gousse pouvant avoir plusieurs allures différentes. Par exemple, la luzerne va donner des gousses enroulées sur elles-mêmes.
La cacahuète (Arachis hypogea), à maturité, montre un photropisme négatif qui fait que le pédoncule se rapproche du sol et enfonce la gousse dans la terre. Le trèfle (Trifolium) a une gousse minuscule. Le genet pour sa dissémination, pratique une ouverture violente de la gousse (autochorie).
Les Faboïdeae ont des rôles alimentaires importants :
- Arachis ? huile et cacahuète.
- Cicer arietinum : le pois chiche.
- Lens culinaris : les lentilles.
- Phasealus : les haricots.
- Pisum sativum : les pois.
- Vicia faba : les fèves.
Elles peuvent aussi avoir un rôle pour les fourrages : Trifolium (le trèfle), Medilago (la luzerne). Elles sont aussi utilisées pour les teintures avec Indigofera (l’indigo) et ont un rôle écologique en formant les landes à genets et à ajoncs.
• Les Cesalpinoideae.
La préfloraison est carénale et l’on a dix étamines libres.
Ce sont généralement des arbres, avec quelques arbustes. Au niveau de cette sous-famille, on a :
- Cercis siliquastrum : arbre de Judée. Cet arbre fleurit avant d’avoir des feuilles. Ces dernières ont leur pétale supérieur interne, contrairement aux papillonaceae. Les étamines sont libres.
- Cassia gloditschia : il peut former de grands arbres avec des épines regroupées par trois au niveau du tronc.
Le caroubier (Ceratonia silique) forme des gousses que l’on appelle « caroube ». Ce végétal est une espèce thermo-méditerranéen (de basse altitude).
4 Les Rosales.
Famille des Rosaceae.
Dans ce groupe, on trouve les fraisiers, les cerisiers… Ils peuvent être des arbres, arbustes ou des herbacés. Les feuilles sont alternes avec présence de stipules. La fleur est généralement cyclique, basée sur le type 5 (pentamère). Celle-ci est hétéroclamydée : pétales et sépales sont différenciés. C’est une fleur dialypétale, polystémone (nombreuses étamines).
L’ovaire peut être libre ou soudé au réceptacle. Les différentes sous-famille sont : les Rosoideae, les Prunoideae, les Spiracoideae, les Maloideae.
Les Rosoideae ont de nombreux carpelles avec un ovule. On distingue trois cas. Les carpelles sont portés par un thalamus : c’est le cas de la fraise. Chez Rubus, la ronce, on a un certain nombre de petites drupes. Le réceptacle s’est retourné et la partie basale de la fleur entoure les carpelles (périgyne) : c’est le cas de Rosa.
Les Prunoideae ont un carpelle unique (prunier, cerisier).
Les Spiracoideae ont cinq carpelles et les fruits sont appelés « follicules ». Ce sont des fruits secs déhiscents).
Les Maloideae sont caractérisés par un ovaire infère soudé au réceptacle (pommier, poirier).
On peut distinguer une tendance à l’apparition d’un ovaire infère et à la réduction du gynécée. C’est une famille majeure d’un point de vue alimentaire : Malus (pommier), Cytonia vulgaris (cognassier), Prunus avium (cerisier), Pyrus communis (poirier), Fragaria vesca (fraisier).
Ces Rosaceae contiennent un certain nombre d’hétérosides cyanogéniques (avec du cyanure) et sont, pour certaines, toxiques (notamment les noyaux mais aussi certains fruits).
Les Rosaceae ont aussi un rôle écologique important, puisque, un certains nombre d’espèce sont pionnières dans l’héliophilie (par exemple le Prunellier [Prunus spinosa] qui supporte la lumière directe).
Sorbus latifolia est une des seules espèces arborées protégée. Cette plante est intéressante car elle est un hybride entre deux espèces : Sorbus aria à feuilles non découpées et Sorbus torminalis à feuilles découpées.
5 Les Fagales.
La famille des Fagaceae.
Cette famille comporte trois genres principaux qui constituent les forets françaises : Quercus (chêne), Fugus (Hêtre), Castanea (Châtaignier).
Les Fagaceae appartiennent au groupe des arbres à chatons ou amentifères. On dit cela car les inflorescences males constituent des chatons.
Les espèces sont monoïques.
Si l’on regarde un chaton male de châtaignier, on observe un épi de cymes de fleurs. Ce chaton possède un nombre constant d’éléments qui chacun donne une cyme bipare. Toutes ces cymes sont réduites à une seule fleur ; cette dernière étant elle-même très réduite.
La différenciation des trois genres principaux se fait au niveau des fleurs males et du fruit qui est toujours un akène monosperme.
Le châtaignier donne trois akènes monospermes car la cyme femelle possède trois fleurs. Chez le hêtre, on trouve deux akènes. Le chêne ne porte lui, qu’un akène car il n’y a qu’une seule fleur au niveau de la cyme femelle.
Ces akènes sont enveloppés par des bractées ligneuses soudées entre elles. Ces dernières constituent un organe : la cupule ; ce qui explique le nom de « cupulifère » donné à ces fagaceae.
On observe de nombreuses espèces de chênes différents. En France, on en a environ sept espèces importantes au niveau écologique qui se répartissent en deux groupes : les chênes à feuillage caduque et les chêne à feuillage persistant (sempervirent). Les quercus caduques sont composés du chêne pédonculé et du chêne sessile. Les quercus persistant regroupent les chênes verts (Quercus ilex), le chêne liège (Quercus suber) et le chêne kermes (Quercus coccifera).
Le chêne liège est une espèce intéressante : c’est une plante calcifuge (qui fuit le calcaire). On le trouve dans les régions méditerranéennes et sur le littoral atlantique. Ce chêne possède un suber provenant du fonctionnement de l’assise subéro-phélodermique, ce qui constitue une bonne protection contre les incendies. On en fait entre autre, des bouchons pour les bouteilles de vin.
La subériculture : la première opération pour enlever le liège est le démusclage ; c’est comme avec les crêpes, la première est ratée.
6 Les Brassicales.
On note une tendance à la zygomorphie chez les Ibéris.
Les Brassicaceae (Crucifères).
Les inflorescence mime une fleur unique : les pétales externes sont plus grands.
Les crucifères se mangent, notamment les brassica (Napus). Napus esculenta = navet ; Napus oleifera = colza.
Brassica oleaceae : c’est le chou. C’est une espèce spontanée sur les littoraux d’Europe du Nord. Brassica sativus est le radis.
Certaines crucifères ont des propriétés médicinales et elles sont souvent riches en vitamine C : ce sont des anti-scorbutiques.
Certaines crucifères sont ornementales, par exemple, Lunaria biennis aussi appelée, la monnaie du pape.
Cheranthus chereis est la giroflée.
Chez la moutarde, on note la présence de cellules à myrosine : c’est une enzyme qui dégrade les glucosides sulfurés (très présents dans les graines). C’est ce qui donne l’huile de moutarde. En fait, c’est un système de défense.
Lors de la mastication, il y a mélange des enzymes et de l’huile de moutarde, ce qui libère des composés sulfurés au goût de moutarde.
Le pastel (Isatis tinctoria) est utilisé comme plante tinctoriale pour sa couleur bleue. Elle était fortement cultivée au 16ème siècle. Sa chute est due à la découverte de l’indigo (Indigofera sulfructosia). On récupère les feuilles de pastel, on les fait macérer, on les broie puis on en fait des boules : la cocagne.
La rose de Jéricho (Anastatica hierochuntica). Quand ses feuilles tombent, s’il fait sec, la plante se recroqueville en boule ? elle se fait arracher par le vent et est transportée, ce qui est un bon moyen de dissémination.
7 Les Ericales.
La famille des Ericaceae.
Les éricaceae forment la famille des bruyères, de la myrtille. Ce sont des plantes ligneuses, en arbustes dont les feuilles sont très variables ; généralement simples, plus ou moins développées. Elles sont du genre érocoïdes : feuilles en aiguille souvent persistantes.
Les landes à bruyère sont sur les sols pauvres.
La taille réduite des feuilles n’est pas explicable par le manque d’eau, mais peut-être à cause de la pauvreté des sols.
Les fleurs sont pentamères : 5S, 5P, 10E, 5C. C’est une famille gamopétale. Les étamines sont à déhiscence poricide. Certaines étamines ont des petites « cornes », des appendices.
Les Ericaceae sous divisées en quatre tribus :
a Les Rhododendroideae.
La fleur est légèrement zygomorphe avec un ovaire supère. Les étamines sont sans appendice.
Le rhododendron est typique de l’étage sub-alpin. En ce moment, il y a recolonisation de ce milieu.
b Les arbutoïdeae.
Arbutus unedo : l’arbousier. Le fruit est une baie ou drupe. Les feuilles sont bien développées. Arctostaphylos uva ursi est le « raisin d’ours ».
c Les Vaccinoïdeae.
Dans ce groupe, on trouve la myrtille.
L’ovaire est infère ; la feuille développée ; les étamines cornues.
d Les Ericoideae.
Le genre Erica est la bruyère.
Le périanthe est persistant la fleur se dessèche sur l’arbre.
Le fruit est une capsule.
8 Les Lamiales.
Les Lamiaceae.
Les lamiaceae sont des plantes aromatiques.
La tige est carrée (quadrangulaire), les feuilles sont opposées, décussées (perpendiculaire par rapport au niveau de la feuille).
Elles présentent des adaptations aux climats secs (thym, lavande).
On trouve des poils sécréteurs.
Les inflorescences sont généralement en cymes concentrées.
La fleur est : 5Ss, 5P (2 pour la lèvre inférieure et 3 pour la supérieure). La zygomorphie augmente. Ces plantes sont gamopétales, gamopétales.
Chez Ajuga, la lèvre supérieure a disparu.
On a quatre étamines soudées à la corolle. Chez Salvia (la sauge), on trouve un système de basculement qui permet de déposer le pollen sur le dos d’un insecte. On a deux carpelles mais qui donnent un tétrakène : il y a formation de fausses cloisons qui sépare les deux carpelles.
La pollinisation est entomophile.
Dans les lamiales, on trouve : Lavandula, la lavande ; Rosmarinus, le romarin ; Thymus, le thym ; Salvia, la sauge ; Satureia, a sarriette ; Mentha, la menthe.
9 Les Apiales.
Les Apiaceae (anciennement Ombellifères).
Les apiaceae sont généralement herbacées sauf Bupleurum qui est ligneux.
Les feuilles sont composées, découpées (carotte, fenouil) sauf Bupleurum (feuilles simples).
L’inflorescence est en ombelle. On a soit une ombelle simple avec une bractée à la base du pédoncule de chaque fleur, soit une ombelle d’ombellule avec les bractées à la base des pédoncules fixés sur l’ombellule. Les ombelles, en se resserrant, vont imiter une fleur unique.
Chez Seseli, la formule florale est : 5Sl, 5Pl, 5E, 2C.
Le fruit est un schizocarpe (= 2 méricarpes) : exemples, graine de cumin, de carvi.
Exemples de plantes alimentaires :
Anethum graveolens, l’aneth ; Antriscus cerofolium, le cerfeuil ; Coriandrum sativum, la coriandre ; Cuminum cyminum, le cumin ; Daucus carota, la carotte ; Foeniculum vulgare, le fenouil.
Exemples de plantes dangereuses :
Conium maculatum, la grande ciguë ; Cicuta virosa, la ciguë aquatique.
10 Les Astérales.
Les Asteraceae (anciennement Composées).
L’inflorescence est en capitule. Les fleurs sont insérées sur un réceptacle. Le tout simule une fleur unique. A la base de celle-ci, on a un involucre de bractées comme chez les apiaceae.
L’ovaire est nettement infère. Le fruit est un akène, souvent surmonté par un papus (ensemble de poils aidant à la dissémination de la graine par le vent).
Ce sont des plantes à fort pouvoir de colonisation.
Exemple d’Asteraceae :
Cichorium endiva, l’endive ; Lactuca sativa, la laitue ; Taraxacum officinale, le pissenlit ; Helianthus annuus, le tournesol ; Cynara scolymus, l’artichaut ; Artemisia absinthium, l’absinthe ; Artemisia glacialis, le genépi ; Arnica montana, l’arnica.
a Les tubuliflores.
Les fleurs sont actinomorphes, tubuleuses. Exemple : le bleuet, Centauver.
Les anthères sont soudées et forment un tube d’où émerge le style et deux stigmates.
b Les liguliflores.
La fleur est ligulée, zygomorphe, avec cinq dents.
c Les radiées.
Les fleurs sont ligulées avec trois dents périphériques. Au centre, elles sont tubuleuses. On trouve chez les radiées, la marguerite, le chrysanthème.
C’est la meilleure simulation de fleur unique.
Partie 2 :
Structures et modalités adaptatives des cormophytes.
I Introduction.
Une adaptation est différent d’un accommodation.
Quand des plantes se maintiennent dans un biotope donné, on considère qu’elles ont toutes les réponses face à ce milieu : elles y sont adaptées. Il faut toutefois éviter toutes les interprétations finalistes : les plantes n’ont pas de conscience. Tout vient de la concurrence, de la compétitivité. Certaines plantes répondent mieux à certains milieux que d’autres. Ces adaptations sont au niveau héréditaires (niveau génotypique).Adaptation : propriété héréditaire, physiologique, anatomique ou morphologique permettant à une espèce de se maintenir et de se développer dans des conditions écologiques particulières.
L’adaptation est plus ou moins stricte.
Exemple du chêne vert. Le feuillage est persistant ; les feuilles petites, dures. C’est une espèce sclérophylle, sempervirente. Il se développe bien sous climat méditerranéen (avec période sèche estivale). Ce chêne n’est pas exclusif à ce climat. On le trouve aussi en climat océanique.Les facteurs qui jouent le plus sont les facteurs de compétition entre espèces. Les plantes les plus adaptées à un milieu seront les plus compétitives. Accommodation : Pour une même espèce, avec deux pieds plantés à des endroits différents (plaine/montagne), on aura des plantes à morphologie différente. La graine n’est pas touchée ? les modifications touchent le niveau phénotypique.
Définition : une accommodation est une modification réversible des caractères anatomiques, physiologiques d’un organisme selon les variations des facteurs du milieu.Dans la Sierra Nevada espagnole, au sud de Grenade, en montagne sèche, quand on passe de 1500 mètres à 3000 mètres, on retrouve des genets purgatifs (papillonacées). A 1500 mètres, ils sont érigés (1,5 à 3 mètres), à 2500 mètres, ils sont beaucoup plus petits et à 3000 mètres, ils sont un port en boule comme des xérophytes épineuses en coussinet.
II Les rythmes biologiques.
A Mode de végétation.
Le plus important est de voir le mode de végétation et la capacité à fleurir.
1 Les plantes à une floraison.
a Plantes annuelles (thérophytes).
Exemples : le blé, le coquelicot.
La graine donne une tige et une(des) fleur(s) puis des graines et un pied qui meurt.
b Plantes bisannuelles.
La première année, ces plantes forment l’appareil végétatif qui accumule des réserves. La seconde année, il y a utilisation des réserves pour fleurir, fructifier puis mourir.
Exemples : Oignon (Allium cepa), la cardère ou cabaret des oiseaux (Dipsacus sylvestris).
Allium donne une rosette, la cardère donne un capitule.
La cardère était utilisée pour carder la laine.
c Plantes pluriannuelles.
Ce sont des plantes vivaces (vivent 10 à 15 ans) qui ne fleurissent qu’une fois puis meurent. On y trouve l’agave (Agave americana).
Ces plantes sont aussi appelées espèces monocarpiques.
On trouve aussi la carline à feuilles d’acan (Carlina acanthifolia) qui présente un gros capitule. Elle est acaule (= sans tige). Pendant plusieurs années, il y a fabrication de rosettes de feuilles et mises en réserve dans les racines pivotantes. Cette plante fleurit ensuite puis disparaît.
2 Les plantes à plusieurs floraisons.
On trouve ce cas chez les arbres, les arbustes, les buissons :les plantes ligneuses. On en trouve aussi chez des herbacées avec rhizomes (exemple : Iris. C’est aussi le cas de plante à bulbe comme la tulipe. Ce dernier exemple n’est pas parfait car il se présente d’autres problèmes.
B La phénologie foliaire.
On distingue les feuillages caduques des feuillages persistants.Les feuillages persistants sont souvent présents dans les climats méditerranéen mais aussi en forêts boréales. Le renouvelage des feuilles se fait tous les ans et demande une grande dépense en énergie. Toutefois, les feuilles sont neuves et donc, plus efficaces.
La phénologie est la façon dont se succèdent les étapes de la vie d’une plante.
III Les types écologiques.
Selon la taille d’un végétal, de là où sont les bourgeons, on peut classer les végétaux en grands types biologiques. Ces types ont été définis par Raunkiaer (originaire de l’Europe du Nord).
La neige est une protection pour les bourgeons contre le gel. Ces plantes ainsi protégées, vont différer des non-protégées. On a donc des différences qui permettent de classer ces végétaux en divers types écologiques.
On prend en compte la bonne saison et la mauvaise saison selon les différents lieu du monde.
- Si les bourgeons sont à plus de 50 centimètres, on parle de Phanérophytes.
- Les chaméphytes (comme la myrtille) sont des végétaux ligneux, vivaces dont la hauteur n’excède pas 50 centimètres.
- Les hémicrophytes (comme le pissenlit, les orties) ont des bourgeons souvent au ras du sol et ont un système de plante en rosette.
- Les géophytes ont des bourgeons de rénovation dans le sol (bulbe, rhizome, tubercule). Ils ont aussi souvent de gros organes de réserve.
- Les thérophytes sont des plantes annuelles, sans bourgeon, qui persistent à l’état de graine.Il a été montré que la période de développement de la plante est variable. Elle est fonction des conditions écologiques du milieu. Pour une plante saharienne, de la germination à la fructification, il ne se passe qu’entre 8 et 15 jours. Ces plantes sont dites éphémérophytes. Elles ont des feuilles naines et de petites fleurs. Elles sont parfaitement adaptées au milieu saharien. (absence de pluie ou pluie localisée dans le temps). Une averse de 4 à 5mm permet le développement de la plante.
Une autre classification est basée à partir du premier bouton floral jusqu’au premier fruit fertile. Si l’intervalle de temps est inférieur à 20 jours, on a des plantes tachythérophytes. Entre 20 et 30 jours, ce sont des sténothérophytes. Entre 30 et 50 jours, ce sont des mésothérophytes. S’il se passe plus de 50 jours, ce sont des eurythérophtes.
A partir des types biologiques, dans un milieu donnée, on définit des spectres biologiques. Ces spectres sont donc variables en fonction du milieu et chaque milieu aura une ou deux espèces dominantes.
IV Multiplication et dissémination sous forme végétative.
Les plantes issues de la multiplication végétative ont le même génotype : elles perpétuent les caractères de la plante mère. Il n’y a pas de brassage génétique. Les individus sont tous des clones.
Pour les plantes dioïques, tous les clones ont le même sexe.
Par exemple, l’Elodée du Canada, en France, b’est présente que sous forme femelle, qui se reproduit donc par multiplication végétative.
A Multiplication par organes non spécialisés.
1 Marcottage naturel.
Exemple de la ronce (Rubus).
On a isolement d’une partie d’un végétal en enracinement. Une tige se courbe en anneau et entre en contact avec le sol : il y a formation de racines adventives qui fixent la tige au sol. Après la coupure, on a un nouvel individu, isolé, identique au pied mère.
Le marcottage existe aussi chez les arbres.
Exemple de l’Epicéa des Alpes, dans les dolomites Italiennes.
Les branches basses sont plaquées au sol par la neige. Elles vont s’enraciner et donner un nouvel Epicéa en aval du premier. Le nouvel individu se séparera de la plante mère. Ce phénomène peut se répéter de nombreuses fois.
On trouve la même chose pour le rhododendron dans les Pyrénées.
2 Le bouturage.
Les racines se forment après séparation (naturelle) de la plante mère. A l’état naturel, on observe ce phénomène chez les figuier de barbarie, chez Sédum (crassulaceae).
? La nouvelle plante est toujours un clone de la plante mère.
B Multiplication par organes spécialisés.
1 Les stolons.
Exemple du fraisier.
Les stolons sont des rameaux spécialisés, à croissance horizontale dont les entre-nœuds sont fortement étirés et portent des feuilles très courtes, réduites à des écailles. Les stolons permettent l’exploration du milieu puis la prise de racine a lieu plus loin.
2 Les racines drageonnantes (ou drageons).
Ce sont des racines généralement horizontales sur lesquelles apparaissent les bourgeons. Exemples : framboisier, peuplier.
3 Les organes de réserve.
Les organes de réserve jouent un rôle de réserve mais aussi de dissémination.
a Les tubercules.
• Les tubercules racinaires.
Des racines adventives vont se tubériser (se renfler de réserves). Exemple : le dahlia dont les racines se renflent et accumulent de l’inuline. Il y aura séparation des tubercules et donc, autant de nouvelles plantes que de tubercules. Exemple : la ficaire (Ranunculus ficaria) a des tubercules avec amidon. Elle possède aussi des bulbilles permettant la dissémination.
• Les tubercules caulinaires.
Exemple : la pomme de terre (Solanum tuberosum). Les tubercules ont un rôle de réserve et de dissémination. On trouve aussi les topinambours (Heliantus tuberosus) et Stachystubifera, le crosne du Japon.
b Les bulbilles.
Les bulbilles sont aussi des organes de réserve. Ce sont des petites bulbes apparaissant à l’aisselle d’une feuille ou d’une inflorescence.
Exemple : le lys (Lilium bulbifera) possède des bulbes qui se détachent, ce qui permet la dissémination de la plante
Ces organes existent aussi chez la ficaire où les bulbilles restent dormantes si elles ne se détachent pas du pied mère.
Chez certains ails, les bulbilles sont entre les fleurs, au niveau de l’inflorescence. Allium est un genre de Liliaceae.
Les saxifragaceae sont des plantes rupicoles (qui poussent sur les rochers) où la base de la tige montre des bulbilles rougeâtres pouvant assurer la multiplication asexuée.
Les bulbilles ou bourgeons épiphylles (Bryophyllum) ont des bourgeons qui se répartissent tout autour de la feuille. Ils vont se détacher et donner de nouveaux individus. Ils n’ont pas de réserves (ce ne sont pas de vraies bulbilles).
V Reproduction sexuée et stratégie de pollinisation.
- Emission de grains de pollen.
- Transport du pollen et dépôt sur les stigmates (fleur de même espèce).
- Développement du tube pollinique, une fois le pollen déposé.
A La répartition des sexes.
Les fleurs sont le plus souvent hermaphrodites (à 75%) mais on trouve aussi des fleurs monoïques ou dioïques.
Le frêne (Fraxinus excelsior) possède des samares (fruits) servant à la dissémination des graines. Dans une population de frênes, on trouve 40% de mâles, 4% de femelles et 56% d’hermaphrodites (dans une population d’une région donnée, étudiée). ? Cet arbre a la potentialité d’être hermaphrodite.
On peut définir trois grands types de pollinisation :
- Autogamie directe :
Le grain de pollen féconde la fleur d’où il vient. Il y a autofécondation. Le transport est court.
- Autogamie indirecte :
Le grain de pollen va féconder une fleur du pied ou bien une fleur d’un clone de la plante dont il est issu. L’autogamie indirecte est aussi appelée Geitonogamie. Cette pollinisation existe chez les hermaphrodites et les monoïques.
- Allogamie :
Le grain de pollen va féconder la fleur d’un individu génétiquement. La pollinisation est croisée. Cette pollinisation existe chez les espèces monoïques ou dioïques.
Il existe des cas où l’autogamie est obligatoire. Par exemple, chez la violette odorante (Viola Odorata), on trouve deux types de fleurs. Les fleurs de printemps s’ouvrent (corolle ouverte) et permettent tous les cas de pollinisation. Les fleurs d’été ne s’ouvrent jamais : ce sont des fleurs cléistogames ne présentant qu’une pollinisation par autogamie directe.
Ce phénomène existe aussi chez Oxalis acetosella.
Le paradoxe : Les deux sexes sont rapprochés, ce qui laisse penser que l’autogamie est de règle. En fait c’est toujours l’allogamie qui domine.
L’allogamie est créatrice de diversité, augmente le brassage génétique ? elle accroît les capacités d’adaptation des espèces.
L’autogamie diminue, elle, les capacités d’adaptation par diminution du brassage.
Il existe des pressions de consanguinité.
Pour les espèces cultivées ou sélectionnées, il faut diminuer, le plus possible, le brassage génétique. Chez beaucoup de plantes principalement allogames, l’autogamie peut intervenir en dernier recours (s’il n’y a pas de pollinisation croisée) pour assurer la reproduction.
Exemple de la stratégie du rhododendron (Ferrugineum), une éricaceae. C’est une plante entomogame. Elles ont 10 étamines dont 5 grandes et autant de petites.
La pollinisation, par un système de tulle montre que la plante est autogame (aucun échange de pollen avec l’extérieur n’est possible). Si l’on refait la même expérience mais en enlevant les petites étamines, on voit que l’on n’obtient jamais de graine. L’autogamie est donc réalisée à partir des petites étamines ; les grandes servent à la pollinisation croisée.
L’autogamie est un système de « roue de secours », une adaptation aux conditions.
Pour diminuer l’autogamie, il y a décalage temporaire de la maturation des anthères et des stigmates (dichogamie). On trouve deux cas : la protandrie, les anthères sont mures avant les stigmates (tournesol) ; la progynie, la partie femelle est mure avant la partie mâle (exemple, l’arum).
Exemple de l’avocatier (Persea gratissima).
Le développement de l’avocatier se fait sur deux jours.

Il y a obligation de pollinisation croisée. Les individus sont soit protandres, soit protogynes.
Dans une même fleur, il a souvent séparation spatiale maximale des étamines et des stigmates. Par exemple, les étamines, dans de nombreux cas, sont beaucoup plus grandes ou petites que les stigmates ? C’est l’erchogamie.
Le plus efficace pour empêcher l’autogamie est une incompatibilité génétique entre le pollen et les stigmates (comme chez les primevère). Chez la primevère, on note la présence de deux types de fleurs : fleurs à étamines longues et styles courts ; fleurs à étamines courtes et à styles longs. On a dans ce cas, erchogamie et auto-incompatibilité.
Les allocations de ressources.
Quand on analyse les comportements de végétaux ou d’animaux, on observe en terme de coût (énergie, ressources…).
L’autogamie demande peu de ressources pour les grains de pollen (peu de grains avec peu de ressources), donc, un faible investissement. On verra chez ces mêmes espèces, une augmentation des ressources des ovules et des graines.
La primevère (Primula farinosa), allogame stricte, a été étudiée et comparée à trois autres espèces autogames. ? Les trois autogames produisent 5 à 10 fois moins de pollen mais deux fois plus d’ovules. On peut faire le rapport P/O (pollen fabriqué/Ovaires fabriqués). Le P/O de farinosa est de 2000 environ et chez les autres, il n’est que de l’ordre de 200.
Il a aussi été montré que la corolle des autogames était plus petite que celle des autres.
Une espèce allogame a un P/O ? 6000 ; un autogame a un P/O ? 30 et une espèce clestogame a un P/O ? 5.
B La Pollinisation.
1 L’anémogamie.
L’anémogamie concerne 20% des espèces florales françaises dont les graminées (les Poaceae). Il faut que le pollen soit de petite taille (10 à 30µm), léger et transportable par le vent. De plus, les plantes « pratiquant » l’anémogamie ne doivent pas regarder sur la quantité de pollen produit.
Par exemple, un pied de maïs donne 50 millions de grains de pollen et seul 1 grain sur 50000 sera utilisé. Le noisetier porte des chatons qui libèrent chacun, 4 millions de grains de pollen par jour. Les chatons se développent avant les feuilles car ces feuilles feraient obstacle au pollen.
Chez les Poaceae, le pollen est fabriqué par des étamines à filet long et mobile.
Pour les espèces anémogames, les fleurs femelles possèdent des stigmates collants (noyer) ou plumeux et longs (Poaceae).
Les fleurs mâles et femelles sont habituellement, relativement simples. Le périanthe, peu développé. Certaines espèces sont même dans pétale, comme les chênes et les orties.
2 La zoogamie.
Dans la zoogamie, le vecteur est un animal, souvent un arthropode. Dans la grande majorité des cas, ce sont des insectes et l’on parle d’entomogamie. D’autres animaux peuvent aussi intervenir, comme le colibri. Les baobabs (Andosonia) sont pollinisés par une chauve-souris : les fleurs s’ouvrent donc à la tombée de la nuit.
Dans le cas des interactions plantes/insectes, le phénomène peut aller jusqu’à une co-évolution : un seul type d’insecte pourra polliniser un seul type de plante.
Les stratégies d’attraction :
• Les signaux optiques.
Les pétales peuvent être vivement colorés et chez les liliaceae, même les sépales sont pétaloïdes. Le développement des bractées joue aussi un rôle : chez l’arum, une grande bractée se développe et piège les insectes.
Les étamines aussi peuvent être colorées.
Chez les orchideae, il y a développement d’une corolle représentant un insecte : c’est un leurre.
Chez les sauge, il se développe une « piste d’atterrissage » pour les insectes.
• Les signaux olfactifs.
Les Lonicera periclymenum sont des fleurs surtout parfumées la nuit car elles sont pollinisées par des papillons (Sphinx). La rafflésie donne une fleur d’environ un mètre de diamètre à odeur de viande pourrie (se trouve vers Sumatra). L’Orchis bouc dégage une odeur de bouc. Certaines Ophrys émettent des phéromones sexuelles d’abeilles.
L’arum émet une odeur renforcée par émission de chaleur (+35/40°C).
• Les apports nutritifs.
Les insectes recherchent de la nourriture. Cela peut être du pollen (comme au début de l’évolution) ou des nectaires (glandes synthétisant du nectar) situés à la base des pétales ou des étamines. La production de nectar a aussi un coût énergétique.
• Le groupement des fleurs.
Ce groupement donne une inflorescence plus ou moins compacte qui permet d’augmenter l’attirance envers les insectes.
Le cas des composées ; le capitule de marguerite.
A l’extérieur, les fleurs sont ligulées et souvent stériles et jouent un rôle dans l’attraction des insectes. Les fleurs en tube, internes, jouent un rôle dans la reproduction.
Le problème est que cette compaction favorise la géitonogamie (les étamines d’une fleur vont féconder les fleurs du même pied).
La digitale (Digitalis purpurea), une scrophulariaceae a des fleurs qui se développent de bas en haut. De plus, ces fleurs sont protandres. La quantité énergétique nu nectar diminue en allant du bas vers le haut. Les insectes vont d’abord aller voir les fleurs du bas puis remonter.
Le cas des orchidaceae (monocotylédones).
La fleur présente trois tépales dont une plus développée que les autres (le labelle) qui est souvent mimétique. Le labelle peut être poursuivit en arrière par un éperon.
Les étamines, le style et les stigmates sont soudés en un organe : le gymnostème. Le pollen est agglutiné au niveau de deux pollinies.
L’ovaire est infère, uniloculaire, constitué de trois carpelles.
Sur les trois stigmates, deux sont fertiles et un forme une petite structure particulière : le rostellum. Ce dernier est situé entre les pollinies et les stigmates fertiles. Il a séparation de l’appareil mâle et femelle dans une même fleur : c’est une erchogamie qui fait diminuer l’auto-pollinisation.
La fécondation est réalisée par les insectes. L’insecte va se poser sur le labelle et les pollinies vont se coller sur la tête de l’insecte.
Exemple de la vanille (orchidée) :
Vanillia planiflora. Cette plante est originaire d’Amérique du sud et d’Amérique centrale. Dans ces milieux, elle est pollinisée par un hyménoptère (Melipona), spécifique de la vanille. Cette vanille a été introduite dans d’autres milieux où elle est restée stérile car il n’y avait pas l’insecte. Il a donc été mis au point une pollinisation artificielle.
Exemple de l’orchidée malgache (Angecum sesquipedale).
En 1860, Darwin observe un éperon nectarifère d’une trentaine de centimètres. 40 ans plus tard, on a découvert un paillon nocturne physiquement adapté à la pollinisation de cette orchidée. ? Il y a un mutualisme entre une espèce de plante et une espèce d’insecte (mutualisme exclusif).
La Pollinisation chez les Arum (Arum maculatum).
Dans la région Midi-Pyrénées, on trouve plutôt l’espèce Arum italicum. L’inflorescence est particulière : une bractée (la spathe) et le spadice. Au niveau de la zone d’étranglement de la spathe sort la massue (partie stérile de l’inflorescence).
Dans la partie basse, on trouve des fleurs femelles fertiles ; un peu plus haut, on a les fleurs mâles fertiles. Dans la partie supérieure, des fleurs stériles bouchent l’entrée.
Le spadice va émettre une odeur nauséabonde (pour nous). La massue s’échauffe (facilement + 15°C par rapport à la température extérieure). Cette augmentation de température accroît l’odeur et attire des diptères. Ces insectes arrivent, entrent et fécondent les fleurs femelles. Après 24 heures, les étamines s’ouvrent, en même temps, les fleurs stériles se fanent. L’ouverture permet la libération diptères, chargés de pollen. La spathe libère des sécrétions pour la nutrition des insectes.
Le figuier, Ficus carica, Moraceae.
La fécondation de ce figuier est complexe et a été bien étudiée.
L’inflorescence du figuier est particulière : clinanthe. C’est un réceptacle replié sur lui-même et qui possède plusieurs centaines de fleurs sur les parois. Un diptère du genre Blastophaga assure la pollinisation.
On trouve trois types de réceptacles selon les saisons et deux générations de mouches. Les réceptacles diffèrent de part le type de fleurs présentes à l’intérieur.
Les femelles de diptères passent dans les fleurs de type B. Elles pondent des œufs dans les fleurs femelles stériles. Les embryons se développent et donnent mâles et femelles. Ils s’accouplent aussitôt après l’éclosion (comme dans « loftstory », cf. le prof.). Les nouvelles femelles sont jusqu’aux réceptacles des fleurs mâles B.
En C, le femelles ne peuvent pas pondre car les fleurs femelles fertiles ont de longs styles. Elle déposent uniquement le pollen et ressortent pondre dans les réceptacles A dans des fleurs stériles. En A, les mouches sont sous forme de larves. En mars, il y a éclosion puis accouplement. Les femelles vont pondre en B et la boucle est bouclée.
La primevère (Primula).
La primevère présente des fleurs brévistylées (à gros pollen et papilles peu développées) ou des fleurs longistylées (à petit pollen à papilles bien développées).
Les grains de pollen d’une même fleur a du mal à polliniser cette même fleur à cause des papilles. Il y a aussi incompatibilité génétique.
La sauge (Salvia).
Les sauges ont, au niveau du périanthe, une expansion qui permet l’atterrissage des insectes. La fleur est protandre. L’insecte va appuyer sur la « palette », ce qui fait se replier l’étamine sur l’insecte. Il va sur la fleur au stade au stade femelle, le style et les stigmates sont au niveau du dos des insectes, il y a récolte de pollen. Il y a très peu d’exclusivité pour les insectes.
3 L’hydrogamie.
L’hydrogamie est un cas relativement rare car le pollen est peu résistant à l’eau. Chez les hydrophytes (immergées en permanence), les grains de pollen ont des substances collantes pour pouvoir être captés. Ils sont souvent en forme de fuseau.
4 La répartition des différents types de gamie.
On a environ 10% d’anémogamie (20% en France). Les steppes herbeuses à graminées en ont beaucoup plus.
Les forets fermées utiliseront plutôt l’entomogamie.
IV Conservation et dissémination des graines.
La graine est un organe de conservation au stade diploïde. Elle est le résultat de la transformation de l’ovule après la fécondation. Elle contient l’embryon avec la radicule (donnera les racines), la gemmule (donnera la tige), le ou les cotylédons (donnent les premières feuilles, plus ou moins modifiées) et une coque protectrice (tégument). On trouvera aussi des réserves qui peuvent être dans l’albumen, dans l’embryon ou dans le nucelle.
A Durée de conservation de la graine.
1 Les graines macrobiontiques.
Ces graines peuvent germer après des dizaines ou des centaines d’années. Une graine de lotus a germé après 250 ans. Dans ces cas, les téguments sont durs, épais et résistants.
2 Les graines mésobiontiques.
Elles peuvent attendre 3 à 10 ans après leur formation. C’est le cas des céréales, comme le blé.
3 Les graines microbiontiques.
Ce sont des graines qui ne vivent que quelques jours à quelques mois.
Le cas le plus extrême est le cas des plantes vivipares : la graine germe dans le fruit encore accroché à la plante mère.
Exemple des palétuviers de mangrove.
Les mangroves sont les forêt tropicales d’estuaire ou de bord de mer. Ce sont des formations soumises au balancement des marées. On trouve peu d’espèces arborées.
Après fécondation et si à marée basse, la plantule tombe dans la vase et se fixe immédiatement. Elle n’a plus qu’à se développer. La plantule peut aussi tomber dans l’eau (à marée haute) : elle va permettre la dissémination.
Dans la région, on trouve Poa bulbosa qui est une espèce vivipare.
Les fruits secs (akènes) offrent une bonne protection à la graine. Les drupes, fruits charnus avec noyau en offrent aussi une.
B La dissémination de la graine ou du fruit.
1 L’autochorie.
Ce sont des plantes qui disséminent leurs graines en les expulsant plus ou moins loin.
On peut citer :
- Les fruits turgescents qui éclatent au moindre frottement (capsules d’impatience d’oxalis).
- Les gousses de fabaceae qui, lorsqu’elles se dessèchent, se tordent et s’ouvrent en expulsant les graines.
- On trouve aussi les légumineuses méditerranéennes qui expulsent les graines au moment des incendies.
- Les plantes comme Arachis, à phototropisme négatif, qui font s’enfoncer la gousse dans le sol, par le pédoncule.
2 L’hydrochorie.
Cette dissémination demande une résistance à l’eau.
Cas de l’iris (Iris pseudocorus) : il fait de grandes fleurs jeunes et les graines sont cireuses, ce qui les rend imperméables.
La noix de coco peut flotter sur des milliers de kilomètres.
3 L’anémochorie.
Les graines sont de petite taille (exemple : orchidées).
Les fruits sont renflés en vessie (Baguenaudier). Chez les papillonacées, la gousse forme une vessie remplie d’air.
Les graines portent des plumes, des aigrettes. Exemple : les graines de peuplier, de saule.
Pappus des akènes (pissenlit).
Graines à ailes : samares de frêne.
Il y a un gaspillage important, donc, besoin d’une forte production pour compenser.
4 La zoochorie.
Les vertébrés ont un rôle prépondérant dans cette dissémination. Il y a production de fruits et de graines riches en nutriments, ce qui représente un coût énergétique payé par les végétaux pour attirer les animaux.
a L’exozoochorie.
Il y a dissémination par fixation des graines sur les poils, les chaussettes : dissémination passive sans avantage pour l’animal. L’accrochement peut se faire par des glochidies qui sont comme des harpons, des épines.
b La synzoochorie.
Il y a récolte et stockage pour la consommation ultérieure. Les graines vont être transportées (par des fourmis) (élaïosomes : diverticules enrichis en lipides) ou par des écureuils qui cachent leurs glands ou noisettes.
c L’endozoochorie.
L’animal va ingérer le fruit ou la graine. Il faut que la graine soit restituée par régurgitation ou par défécation. Il y a alors séparation des réserves alimentaires pour l’animal et pour la graine sclérifiée ? résistance au broyage ou à la corrosion par les sucs digestifs.
Le transit par le tube digestif devient obligatoire car l’attaque par les sucs ou par les bactéries intestinales va favoriser la germination de la graine.
Etude du Lierre (Hedera hélix), comparaison du taux de germination.
Les graines extraites de baies mûres (directement sur l’arbre) donnent 74% de germination. On remarque que ces graines sont rapidement envahies par des mycéliums.
Si les graines sont extraites de baies vertes (pas mûres), il y a 100% de germination : les baies sont aptes tôt à germer.
Les graines récoltées au sol après ingestion et rejet par les oiseaux donne 94%% de germination, mais, sans développement de mycélium ? les graines sont nettoyées par les sucs, ce qui permet de ne pas avoir de pourrissement.
L’endozoochorie montre aussi des cas de mutualisme : bénéfices réciproques pour les deux partenaires.
Les vertébrés auraient joué un rôle important dans la recolonisation des terres après les glaciations et autres phénomènes.
Après les glaciations, les hêtres ont recolonisé 3000Km en 3000 ans, soit 1 km par an : l’action des animaux est obligatoire.
5 La barochorie.
Les fruits ou graines tombent sous l’action de la gravité : marronnier, chêne, cocofesse (Lodoïcea).
Cette forme de dispersion a une importance relative.
En milieu forestier, on voit qu’il y a une large domination de la dissémination par les animaux.
A leur début, les forêts sont ouvertes et l’anémochorie domine. Au fil du temps, la forêt se referme et le type de pollinisation évolue. Plus la formation est ouverte, plus la pollinisation est réalisée par le vent.
V La fixation des plantes.
La majorité des plantes est fixée dans le milieu souterrain. D’autres, comme les épiphytes, ont une fixation aérienne.
A Le système racinaire.
On a une racine principale, issue de la radicule, et des racines secondaires (2ème ou 3ème ordre). Dans certains cas, on a des racines adventives. En général, elles naissent au niveau de rhizome, mais elles peuvent apparaître sur des tiges aériennes.
Les racines pivotantes : la carotte. Ce sont des expansions verticale avec peu de racines secondaires.
Les racines fasciculées. Il n’y a pas de racine principale dominante : les ramifications sont importantes. Exemple : le hêtre adulte. Les graminées ne sont pas vraiment dans ce groupe car elles ont des racines adventives.
Pour une même espèce, le système racinaire peut changer avec l’age mais aussi, en fonction du type de sol.
En fonction du temps :
Le jeune hêtre a des racines pivotantes pour aller chercher l’eau et les sels minéraux. Au bout de 4 à 5 ans, il développe des racines secondaires. Ensuite, il va mettre en place un système de racines fasciculées.
En fonction du type de sol.
Exemple 1 : Pinus sylvestris.
On trouve deux types de racines : des racines superficielles captant les précipitations et des racines pivotantes qui sont en contact avec la nappe phréatique. Si la nappe phréatique descend, il y aura diminution de la croissance de l’arbre.
Exemple 2 : Une graminée, Ligeum spartum.
Ces graminées sont souvent sur des sols avec une croûte gypseuse (le pH augmente). Les racines vont se développer sous la couche sableuse qui se trouve au dessus de la couche dure.
Exemple 3 : Les podzols (sol de la forêt landaise, indifférencié).
L’horizon A2 est appauvri et essentiellement constitué de sable : il ne retient ni l’eau ni les éléments nutritifs.
Dans cet horizon, les racines ne se développement pas du tout. Un système de racines superficielles existe ainsi qu’un système pivotant.
Exemple 4 : Les sols bruns.
Dans les sols bruns, très homogènes, les racines se répartissent uniformément..
B Modes particuliers de stabilisation.
1 Les racines de contrefort.
Elles existent uniquement chez les arbres tropicaux. Elles se développent dans la partie basse du tronc.
2 Les racines échasses.
On les trouve, par exemple, dans les zones de mangrove. Ces racines partent du tronc et plongent dans le sol (Rhizofera). Elles permettent au tronc de se retrouver au dessus du niveau de l’eau quand il y a marée haute.
Les pneumatophores sont des racines aériennes arrivant à la surface du sol et permettent d’alimenter la plante en O2 (Avicennia).
C Les xérophytes, adaptation des racines.
Les xérophytes font des prospections profondes, jusqu’aux nappes phréatiques (? plantes phréatophytes). Exemple : l’Acacia du Sahel, il peut avoir des racines de plusieurs dizaines de mètres de profondeur. Les nappes doivent être bien alimentées en eau, sinon les plantes meurent.
Il peut y avoir enracinement latéral, peu profond, pour profiter de la moindre goutte de pluie (pour les plantes annuelles). En quelques heures, il y a formation de racines de pluie qui disparaîtront en quelques heures.
Le Zygophyllum d’Israël a des racines dormantes qui deviennent fonctionnelles en cas de pluie.
D Fixation en milieu aériens (lianes, épiphytes).
1 Les lianes.
Les lianes sont des plantes à tige longue qui s’appuie sur divers supports (souvent une autre plante) pour élever son feuillage et le reste (recherche de la lumière).
a Système non préhensile (Rosier).
Par circumnutation, la tige, quand elle pousse, a son sommet qui ne reste pas immobile : il y a enroulement autour d’un support, sans organe particulier.
b Système préhensile.
Dans ce cas, il y a développement d’organes particuliers pour s’accrocher.
Par exemple, le lierre développe des racines adhésives.
Pour les vrilles, on distingue deux cas : les vrilles caulinaires où la tige donne les vrilles (exemple : Vitis vinifera) et les vrilles foliaires où les feuilles donnent les vrilles.
Les vrilles vont s’enrouler dans les deux sens, ce qui leur procure une meilleure élasticité et qui rapproche le support da la plante (Bryone : feuille vrille).
Les pois ont une partie de la feuille qui donne la vrille.
Lathyrus aphaca : Il y a apparition de stipules développés qui assurent l’assimilation chlorophyllienne car les feuilles sont transformées en vrille
La salsepareille (Smilax aspera) a des stipules transformés en vrille.
2 Les épiphytes.
Ce sont des plantes vivant sur d’autres végétaux. Ces derniers servent de support sans servir de nourriture à l’hôte (pas de parasitisme). C’est le cas des lichens, des mousses, des fougères. Beaucoup d’orchidées sont épiphytes.
Il n’y a pas de sol, ni de réservoir d’eau. Les arbres supports doivent avoir des aspérités pour que les épiphytes se développent dessus (réserves d’eau, de terre dans les creux de l’écorce).
Les épiphytes xérophytes comme Tillandsia absorbent l’eau par leur tige.
Les plantes carnivores ont des apports nutritionnels par les animaux consommés et les apports azotés viennent aussi des animaux.
Parfois, des lianes deviennent épiphytes: problème de classement.
A l’inverse, des Ficus (épiphytes) développent des racines qui s’insèrent dans le sol : ils deviennent des lianes).
Exemple : le ficus étrangleur du palmier à huile. Les racines descendent le long du tronc et arrivent au sol. Là, il a une bonne alimentation et étrangle la plante hôte qu’il gardera comme support.
Les Algues.
I Evolution et phylogénie.
A Que sont les algues ?
On ne peut pas donner d’image typique des algues.
Les couleurs observées, bien que différentes, ne sont pas spécifiques.
On trouve des algues de quelques micromètres (diatomées) jusqu’à plusieurs mètres. à On a une grande diversité : diversité de taille ; diversité morphologique : les Chlamydomonas sont des unicellulaires, Ectocarpus est filamenteuse, Ulva lactuca est structurée ; diversité écologique : on peut trouver des thallophytes dans les cours d’eau (diatomées, algues vertes, cyanobactéries), dans la neige (algues vertes) et sur terre.
Les laminaires présentent une convergence avec les végétaux supérieurs.
Finalement, on observe de fortes adaptations aux différents milieux.
B Position phylogénétique : systématique et définitions.
Les thallophytes regroupent tout ce qui est différent des cormophytes.
Chez ces thallophytes, une cellule mère va générer des cellules filles emprisonnées dans la paroi de cette cellule mère.
Chez Chara, on trouve des cellules de reproduction sexuée qui préfigurent la graine.
Les Cryptogames ont des structures de reproduction cachées alors que les Phanérogames ont des structures de reproduction visibles.
Les caractères utilisés en phylogénie sont :
- Les caractères ontogéniques : le passage par des phases archaïques.
- Les caractères paléontologiques.
- La morphologie comparée.
- Les caractères moléculaires : ces caractères peuvent être discontinus, continus. On prendra le cas des gènes 16s. Ce dernier est très utilisé pour la mise en place des phylogénies car il est présent chez tous les organismes. Ils sont aussi abondants et très faciles à isoler.
Les algues sont des eucaryotes autotrophes dont l’appareil végétatif est un thalle (sans tissu différencié). On leur rapproche les cyanobactéries et les prochlorobactéries.
1 Ectocarpus siliculosus.
L’appareil reproducteur est un cyste pluriloculaire avec des grains formés par mitoses.
Lorsque l’on n’a qu’une loge, c’est un cyste uniloculaire (on est chez le sporophyte), porté par des cellules contenant plusieurs spores (obtenues par méiose).
Quand on n’a pas de cyste uniloculaire, on se trouve chez le gamétophyte.
2 Sphacelaria plumula.
On trouve une grosse cellule apicale et une tige formée d’une seule rangée de cellules.
Autour de l’axe central, on a un cortex cellulaire.
Les ronds orangés sont des plastes.
Le thalle est uniaxial, polystylé.
3 Fucus.
Fucus est une espèce dioïque. Les points observés à la surface sont des conceptacles. Les très grosses granules sont les parties femelles. Les tas de filaments sans structure épaisse sont la partie mâle.
4 Laminaria digitata.
On trouve deux zones corticales pigmentées (le méristoderme) avec des structures de reproduction (cystes et paraphyses). On trouve ensuite deux zones composées de grosses cellules (les cellules d’assises) et une zone interne avec des filaments (la médullaire).
On trouve un plaste divisé en vésicules, reliées entre elles par une membrane.
II Diversité de thalle et mécanisme de croissance.
A Organisation générale.
Les archéthalles présentent des cellules non unies (indépendantes), isolées ou non.
Les nématothalles montrent des cellules dépendantes avec des thalles filamenteux pouvant évoluer en lame. La croissance va être diffuse ou localisée en région apicale. Les cellules sont reliées par le plasmodesme.
Les cladothalles sont des thalles filamenteux à axe de croissance préférentiel.
1 Les Archéthalles.
a Les cellules les plus simples.
On trouve des cellules indépendantes, isolées ou incluses dans un cénobe. On trouve par exemple, Chlamydomonas qui possède deux flagelles.
b Les cellules isolées.
Dans ces cellules, on trouve : une paroi, un corps intracellulaire, des dyctiosomes, un gros plaste avec une masse protéique (pyrénoïde), un stigma dans le plaste (la structure photosensible qui permet l’orientation des cellules nageuses), deux vacuoles pulsatiles participant à l’osmorégulation, deux équiflagelles. La division a généralement lieu par bipartition.
c Les associations de cellules immobiles.
Chez Scénédesmus, on a quatre cellules qui peuvent se séparer et donner de nouveaux cénobes. Ces cellules sont associées par un ciment.
d Les thalles pluricellulaires.
Dans ce cas, on peut trouver des cyanobactéries a structure pluricellulaire.
Chez Zygnéma, les cellules sont indépendantes, non réunies par un plasmodème.
Chez Pediastrum, il y a association cellulaire. Chaque cellule donnera un cyste qui permettra la formation de spores flagellées restant groupées dans un mucilage. L’appareil végétatif va se transformer et donner un adulte : les centres opérateurs organisent la mise en place du réseau de filaments.
e Les associations cellulaires mobiles.
Chez les chlorophycées, on observe une évolution de ces structures.
De Gonium, groupement de 16 cellules en forme de coupelle, on passe à Pandorina, comprenant toujours 16 cellules mais cette fois, de forme sphérique. Finalement, on arrive à Eudorina, composée de 64 cellules avec une spécialisation apparente. Le dernier stade est Volvox.
Chez Volvox, composé de plusieurs milliers de cellules, on trouve des petits ronds verts (à l’intérieur) qui sont des cénobes fils.
Cette évolution morphologique va être suivie par une spécialisation des cellules (chez Gonium, toutes les cellules sont indépendantes). Chez Eudorina, de nombreuses cellules ont une fonction reproductrice. La tendance évolutive est à la réduction du nombre de cellules reproductrices.
Chez Volvox, les gonidies (appareils reproducteurs) apparaissent au moment de la reproduction. Ces gonidies sont des retournements de membrane.
2 Les Nématothalles.
Chez les nématothalles, on observe un changement d’organisation fonctionnelle par rapport aux archéthalles.
Ces thalles peuvent évoluer en lame.
Les relations intercellulaires se font par l’intermédiaire d’une synapse, bouchée par un bouchon synaptique
b Les divers types de filaments.
On peut trouver des filaments prostrés, dressés, hétérotriches.
Les filaments haplostiques sont des filaments composées d’une unique file cellulaire (exemple : Ectocarpus).
Les filaments polystiques sont composés de plusieurs rangées de cellules (exemple : Entéromorpha).
Parmi les espèces à thalle siphonné, on trouve Vaucheria.
3 Les cladothalles.
Ces thalles ont une axe préférentiel de croissance : le cladome primaire.
La croissance est continue. Sur chaque cladome, on va trouver des pleuridies. Les pleuridies sont de petits filaments ayant une croissance continue.
b Les différentes structures.
Les cladothalles uniaxiaux ont un axe central composé d’une file de cellules (haplostiques).
Les cladothalles polystiques ont un axe central avec cortex.
Les cladothalles multiaxiaux présentent des articulations entre des articles calcifiés : c’est une zone sous le cortex pleuridien (exemple : Corallina officinalis).
c Variations morphologiques.
Les lames foliacées (exemple : Delesseria).
La lame donne un cladome primaire qui donne lui, des cladomes secondaires et des pleuridies couchées entre les cladomes secondaires.
Les cladomes à filaments siphonnés.
Ils sont trouvés chez les Choloropyceae, Bryopsidales.
Les cladomes à filaments hémisiphonés.
Dans ce cas, on a des articles plurinucléés. Entre les cellules, la paroi vient former un bouchon d’obturation. Des vacuoles internes, riches en polysaccharides, viennent former la paroi.
Les cladomes de Florideophyceae.
Ces cladomes présentent de nombreuses variantes ; ils peuvent être formés de cellules ou de filaments è ce sont des cladomes aplatis.
On trouve aussi des cladomes multiaxiaux : une zone corticale est formée de filaments ou de cellules ainsi que de petits rameaux. Chez certaines algues, le cladome multiaxial disparaît sauf dans la région apicale.
B Mode de croissance.
Toutes les cellules sont aptes à se diviser : Chlorophyceae, Ulothrix, Spyrogyra.
2 La croissance localisée (une zone de croissance).
On peut trouver une zone intercalaire qui est contenue dans l’appareil végétatif.
La croissance trichothallique se fait par des cellules supérieures plus fines et elle est rencontrée dans plusieurs genres d’algues brunes comme chez Ectocarpus.
On peut aussi trouver plusieurs zones de croissance par filament.
3 La croissance apicale.
Cette croissance est réalisée par une cellule apicale et permet la formation de toutes les algues siphonnées. On trouve ce cas chez les Sphacélariales (algues brunes). Si l’on ne trouve qu’une cellule apicale, on aura un cladome uniaxial ; si l’on a plusieurs cellules apicales, le cladome sera multiaxial.
C Les thalles de phaeophyceae.
L’appareil végétatif est très développé.
Chez Dyctiota dichotomia, on a une cellule apicale qui donne deux cellules filles puis deux ramifications qui donneront quatre cellules filles puis quatre ramifications.
Les cellules à la base des cellules apicales sont des cellules filles.
La zone corticale donne des cellules pigmentées. Les grosses cellules sont des cellules de stockage.
Fucus sp.
L’appareil végétatif est un disque basal et est aplati. Il peut donner des vésicules aérifères (pour le soutien).
On ne trouve pas de nervure mais une rainure et une zone fertile apicale (le réceptacle).
A l’extrémité, on trouve des cryptes avec une grosse cellule (cellules apicales) avec cinq plans de mitose. Cette cellule est l’origine d’une couche cellulaire qui sera à l’origine de filaments.
On trouve aussi deux zones corticales (méristoderme) assimilatrices.
On distingue aussi des cellules anguleuses qui forment un cortex de soutien.
Les Laminariales.
Les laminariales ont une longue structure érigée : stype avec une fronde et crampon basal.
On distinguera deux organisations différentes : un stype pérenne et une fronde annuelle. La zone de croissance est intercalaire et l’allongement du stype entraîne la formation d’une nouvelle fronde.
Sur le stype, à l’extérieur, on a le méristoderme qui montre une croissance en épaisseur montrée par des stries annuelles.
On peut voir des petits canaux qui sont des canaux mucifères et des cellules accolées à paroi criblée.
è Cette ligne des laminariales est parallèle à celle qui a donné les végétaux supérieurs.
D Les structures cellulaires particulières.
1 La paroi ou exosquelette.
a Les algues sans paroi.
Ces algues ont un système de rigidification.
Les Cryptophytes (unicellulaires). Il y a formation de plaques qui sont des inclusions de protéines sous le plasmalemme.
Les Dinophyceae ont des armures de plaques protéiques.
Les Euglenophyceae. Chez Phacus, une espèce striée, on peut voir que les stries sont dues à des plaques articulées entre elles sous le plasmalemme. Les Phacus sont capables d’effectuer des mouvements amiboïdes.
Les Chrysophyceae (Emiliana luxleyi). A l’intérieur, on trouvera des écailles de calcite aussi appelées coccolites ou coccosphères.
Chez les Diatomophyceae (Bacillariophyceae), on trouve un frustule siliceux. Ce sont des espèces souvent pélagiques. On trouve deux types de symétrie : radiale (centrique) ou planaire (pennées). L’extérieur est recouvert par deux valves de silice, emboîtées (l’épivalve est supérieure et l’hypovalve est inférieure). Des ceintures connectives vont sceller les valves.
La division se fait quand une ceinture connective glisse ; il y aura division cellulaire et formation de nouvelles valves. Les nouvelles valves seront toujours des hypovalves (par rapport à celles déjà existantes). è Il y a donc une diminution de la taille. Plus la taille diminue, plus la flottabilité régresse. Au bout d’un moment, il va y avoir reproduction sexuée par zoospores, fusion, formation d’un embryon et formation d’une cellule mère.
b La paroi des algues.
Cette paroi est constituée de deux phases : une phase cristalline (squelette de microfibrilles) et une phase amorphe (matrice prédominante).
Les algues rouges et brunes on une phase amorphe importante qui est un réseau de polysaccharides qui forme un mucilage (origine en boite à œufs).
La phase cristalline.
Cette phase est composée de cellulose, de xylème, de mannane, avec selon les algues, un polymère dominant.
Chez les chlorophycées, la phase amorphe est majoritairement composée d’hétéropolysaccharides sulfatés.
Chez les Rhodophyta, on trouve des galactanes, pour résister à l’immersion.
Chez les Phaeophyceae, on trouvera principalement des alginates.
2 Les flagelles.
Un flagelle est composé de 9 associations de tubulines (a et b) avec 250 protéines associées.
On trouve un corpuscule basal qui permet l’insertion du flagelle ainsi qu’une structure intracellulaire.
Comme organisation de l’appareil cinétique on trouve :
- Cellules acrokontées : les flagelles apicaux.
- Cellules isokontées : les flagelles sont égaux.
- Cellules pleurokontées : les flagelles sont latéraux.
- Cellules hétérokontées : les flagelles sont différents.
- Cellules stéphanokontées : les flagelles ont une disposition rayonnante.
Chez les Rhodophyta, les flagelles sont absents ; chez les Chlorophyceae, les flagelles sont isokontés ; les Hétéroconta ont des flagelles hétérokontés.
Le déplacement des cellules flagellées se fait vers le haut (la fuite, vers le bas).
3 Les plastes.
Les formes archéoplastidiées : on trouve un gros plaste focal ou pariétal.
Les formes néoplastidiées : ce sont souvent des structures cladomiennes.
Les formes mésoplastidiées : ce sont des structures intermédiaires (exemple : Chladophora).
Les algues rouges, Bangyophyceae sont archéoplastidiées ; les Florideophyceae sont néoplastidiées.
Les algues brunes, comme Ectocarpus, sont archéoplastidiées.
b Structure.
Les algues vertes ont des chloroplastes qui sont formés de deux membranes et un système interne, les thylacoïdes. Chez les végétaux supérieurs, on a des granas qui sont des empilements de thylacoïdes.
Ces algues comprennent un pyrénoïde (structure protéique interne) qui ne forme pas de grana et qui permet la synthèse d’amidon intra-plastidiale.
Les algues brunes ont des phaeoplastes. Certains plastes comprennent un pyrénoïde en position déjetée. L’enveloppe externe est formée de quatre membranes : deux membranes comme chez les chloroplastes et deux externe de réticulum endoplasmique.
Entre les deux membranes externes et les deux internes, on trouve une structure avec de l’ADN : c’est le nucléomorphe (un noyau atrophié).
Il n’y a jamais de phycobilisomes.
Chez les Dinophyceae, on trouve une enveloppe classique, composée de deux membranes avec une membrane de réticulum.
Souvent, les thylacoïdes sont groupés par trois (en général).
Chez les Cryptophyceae, les thylacoïdes sont groupés par 2. Ceux de Dinophyceae et Euglenophyceae sont souvent non groupés (parfois, sont par trois).
Les rhodoplastes de Rhodophyceae.
Ce sont des pigments rouges hydrosolubles. L’enveloppe plastidiale est formée de deux membranes. Souvent, on a un seul thylacoïde qui peut faire tout le tour du plaste. Il y a absence de pyrénoïde dans les thylacoïdes des algues rouges.
En microscopie électronique, on voit des points de phycobilisomes : se sont des structures protéiques qui recouvrent les photosystèmes sur les thylacoïdes. Ils contiennent de la phyco-érythrine ainsi que de la phycocyanine.
On retrouve ce système sur les cyanobactéries où les thylacoïdes sont chapeautées par des phycobilisomes.
Les Cryptophyceae présente un nucléomorphe avec quatre membranes.
Les algues rouges ont deux membranes.
c Origine évolutive, la théorie de l’endosymbiose.
On trouve une structure unicellulaire eucaryote qui héberge un procaryote symbiotique : ce sont des cyanobactéries dans des glaucocystophytes.
Le symbiote est intégré à la cellule et subit une perte de son système membranaire (algues rouges).
Une cellule eucaryote « avale » une cellule eucaryote endosymbiotique : ce sont les cryptophytes (quatre membranes et un nucléomorphe).
Il y aura une disparition du nucléomorphe : il reste une enveloppe, deux membranes, deux réticulum mais plus de nucléomorphe.
On trouve des plastes primaires (chlorophyceae, rhodophyceae) et des plastes secondaires (algues brunes).
d Les stigmas.
Les stigmas sont présents chez de nombreuses algues flagellées.
Un stigma est une structure composée de caroténoïdes qui selon les espèces est une dépendance d’un plaste ou un plaste dégénéré.
C’est un photosystème qui intervient dans l’orientation : c’est un cache qui permet de générer une zone d’ombre. De plus, une structure photosensible serait à la base du flagelle. La flavine est la vraie structure photosensible réceptive. On observera une variation de dépolarisation du flagelle qui le fait battre ou non.
III La reproduction.
A Multiplication végétative.
1 La bipartition cellulaire.
Les cellules vont se diviser en deux (exemple : Chlamydomonas). La division donne des cellules filles qui assureront la dissémination ou une réunification dans une colonie (exemple : Diatomées).
Cette bipartition n’existe pas chez les algues rouges.
2 La fragmentation du thalle.
Cette fragmentation est fréquente, même chez les pluricellulaires. Celle-ci permet au végétal d’avoir un côté invasif. On trouve par exemple Caulerpa taxifolia qui envahie les posidonies (Posidonia et Zostera sont des graminées marines). Cette caulerpe est toxique pour de nombreux poissons et coquillages et vont donc diminuer la qualité du biotope.
Par exemple, la Sargasse est une algue ayant envahie la côte européenne venue avec des huîtres du Japon.
3 La différenciation de spores.
L’exemple des Sphacélariales :
On trouvera par exemple des propagules qui sont des structures pluricellulaires.
Il pourra y avoir formation de zoospores comme chez Oedogonium (libération de spores stéphanokontés).
B La reproduction sexuée.
1 Les gamètes et types de gamies.
a L’hologamie.
Il va y avoir de deux cellules correspondant à l’appareil végétatif qui fusionnent. C’est le cas des structures unicellulaires.
b La cystogamie.
Il va y avoir fusion de deux cellules sans qu’il n’y ait de différenciation de gamètes (une cellule par individu).
c La planogamie.
Il va y avoir de gamètes flagellés. Gamètes mâles et femelles peuvent avoir la même morphologie (planogamie isogame) ou une morphologie différente (planogamie anisogame).
d L’oogamie.
Dans ce cas, l’oosphère est non flagellée.
e Exemples.
La cystogamie chez Casmarion (algue unicellulaire) : deux cellules s’accolent (en position perpendiculaire) par leur mucilage. Les contenus cellulaires vont fusionner pour former un zygote.
L’hologamie chez Spirogyra. Deux filaments vont fusionner. Tout passe dans un des filaments. Il y a anisogamie car on trouve un donneur et un receveur.
2 Illustrations de quelques cycles.
On trouve deux générations (une haploïde et une diploïde), c’est un cycle digénétique haplodiplophasique. Les générations haploïdes et diploïdes ont la même morphologie (cycle digénétique isomorphe). Il y a planogamie anisogame.
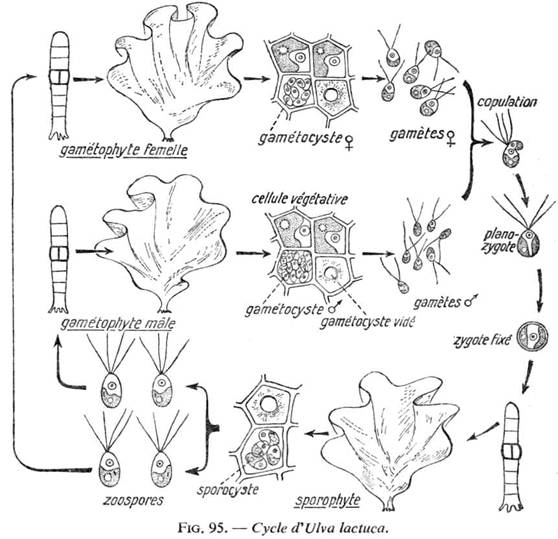
b Codium sp.
Le cycle est monogénétique, diplophasique. La planogamie est anisogame. Les utricules portent des gamétocystes qui donnent des gamètes mâles et femelles qui après fécondation donnent un embryon qui va donner des filaments puis une plante qui porte des utricules.
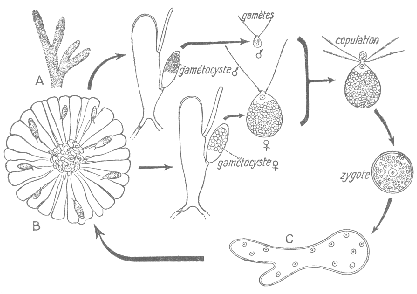
c Chara sp.
Le Chara a un cycle monogénétique et est haplodiplophasique. La fécondation est une oogamie.
La plante présente des gamétocystes mâles (ronds) qui vont libérer par éclatement, des spermatozoïdes biflagellés. Ces spermatozoïdes vont féconder l’oosphère qui reste protégée par une cellule spirale (àgamétange). Il va y avoir réduction chromatique qui aboutit à une cellule à un noyau et à une cellule à trois noyaux. La cellule mononucléée donnera la cellule germinative alors que la cellule trinucléée donnera les réserves en amidon.
Chara pourrait être le chaînon manquant entre les algues et les végétaux supérieurs. « L’œuf » de Chara préfigure la graine car il y a apparition des cellules de protection, d’albumen et d’une cellule germinative.
d Ectocarpus siliculosus.
Ectocarpus présente deux cycles : un cycle digénétique diplohaplophasique et une variante monogénétique diplophasique.
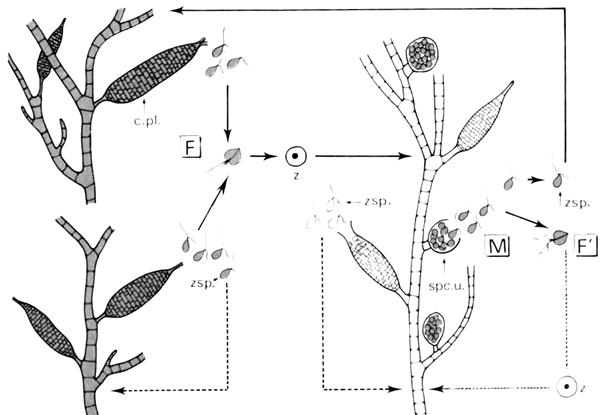
En traits pleins, le trajet digénétique ; en traits discontinus, le trajet monogénétique.
Les gamétophytes haploïdes portent des cystes pluriloculaires qui libèrent des zoospores. Ces zoospores ont deux devenirs possibles : soit, donner un gamétophyte haploïde par germination, soit, participer à la fécondation par planogamie anisogame.
Cette fécondation donne un embryon qui va former un sporophyte diploïde. Sur ce sporophyte, on va trouver deux sortes de cystes.
Les premiers cystes sont les cystes pluriloculaires qui donnent des zoospores à 2N. Ces dernières redonneront directement des sporophytes diploïdes. Les seconds cystes sont les cystes uniloculaires qui libèrent des zoospores à N. Ces zoospores vont alors donner un gamétophyte haploïde. Là, le cycle est bouclé.
On peut avoir une variante ou les zoospores à N jouent le rôle de gamètes et permettent donc la fécondation. Celle-ci donne un embryon évoluant en sporophyte qui pourra redonner des zoospores à N.
e Fucus serratus.
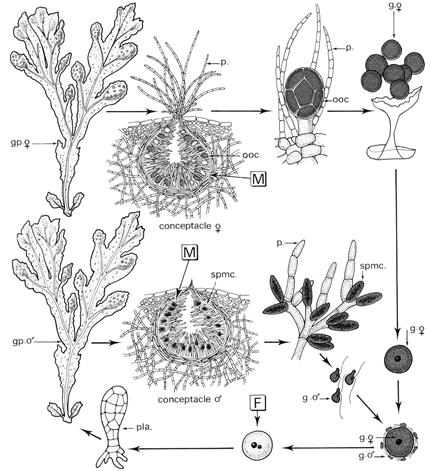
Le cycle de ce Fucus monogénétique, diploïde. La fécondation est une oogamie et les flagelles portés sont au nombre de deux et différents morphologiquement.
f Les Laminaires.
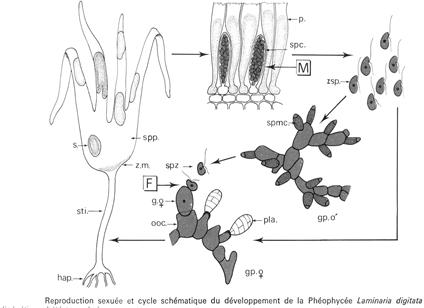
Le cycle présente deux générations, une génération sporophytique et une génération gamétophytique.
Le sporophyte va donner une cellule qui va subir la méiose, donner des cystes uniloculaires. Ces cystes sont insérés entre des cellules stériles, les paraphyses.
Les cystes contiennent des zoospores haploïdes qui donneront des gamétophytes (mâles ou femelles). Ces derniers (les formes mâles) forment des spermatocystes libérant des spermatozoïdes qui vont féconder les oosphères et ainsi, donner un zygote.
è Le cycle est digénétique, haplodiplophasique, hétéromorphe.
g Porphyra umbilicalis (algue rouge).
L’appareil végétatif est un cénobe.
Il n’existe pas de cellules flagellées chez ces algues. On ne trouve donc pas de spermatozoïde mais des spermaties. Ces dernières vont féconder le carpogone qui subit alors trois mitoses et forme huit cellules. Ces huit cellules libérées sont les carpospores.
Elles vont germer et donner un filament incrustant (sporophyte qui se développe sur un support calcaire). Le filament subit la réduction chromatique et donne des spores qui en germant, donneront des lames de Porphyra.
h Polysiphonia elongata (Florideophyceae).
Polysiphonia présente un cycle trigénétique, haplodiplophasique.
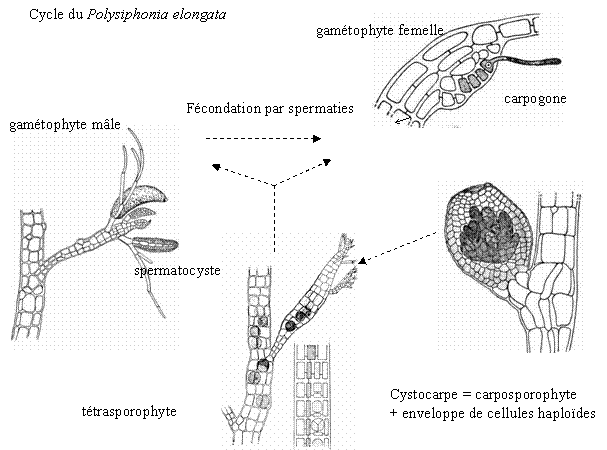
Le gamétophyte mâle libère des spermaties. Les gamétophytes femelles mettent en place des carpogones, surmontés par un trichogyne. Les spermaties vont féconder les carpogones.
La fécondation donne un zygote qui reste fixé sur le pied mère pour effectuer une multiplication cellulaire : il évolue ainsi en carposporophyte. Ce dernier libèrera des carpospores évoluant en (tétra)sporophytes lui-même, donnant des tétrasporocystes, libérant des tétraspores. Ces dernières spores donneront les gamétophytes (mâles et femelles).
Une génération est une structure pluricellulaire venant d’une fécondation ou de la germination d’une spore et qui libère elle-même des spores.
IV Les algues et leur milieu.
A Les types trophiques.
Les milieux de développement des algues sont des zones avec présence de lumière. Le classement se fait selon les sources d’énergie et selon les sources alimentaires.
Les bactéries vertes seront trouvées dans les sources d’eau chaude alors que les bactéries pourpres sont présentes dans les zones soufrées.
On pourra distinguer des organismes à photosynthèse archaïque ou à photosynthèse évoluée.
è On fera une différenciation par rapport à la composition pigmentaire.
La bactériorhodopsine est une protéine permettant de faire toute la chaîne de la photosynthèse à elle seule.
Les plantes effectuant une photosynthèse évoluée comprennent les pigments suivants : fucoxanthine, chlorophylle A, photosystèmes, pigments surnuméraires comme les phycobilines (phycocyanine, phycoérythrine).
Les phycobilines sont surtout présentes chez les cyanobactéries et les rhodophytes.
Le trajet des électrons.
Chez les algues et les cyanobactéries, le principe est le même : l’eau donne l’oxygène qui sera libéré sous forme d’O2 gazeux. Les électrons seront ensuite déplacés.
Les bactéries pourpres et vertes ont un photosystème anoxygénique qui montre un trajet cyclique des électrons.
B Les algues benthiques.
La flore marine est composée à 93,8% d’algues ; 5,5% sont des monocotylédones marins (exemples : Zostères, Posidonie) ; quelques lichens dans les zones de balancement de marées.
2 Les contraintes à la vie marine et adaptations.
On assiste à une raréfaction de la lumière en fonction de la profondeur. Plus la profondeur augmente, plus la quantité et la qualité de la lumière diminuent. Le rouge et le violet disparaissent les premiers. Le vert reste fort longtemps.
On observe donc :
- Des phytoplanctons dans les premiers mètres qui absorbent surtout le rouge et le bleu.
- L’énergie photonique est inversement proportionnelle à la longueur d’onde. (E=h.c/l)
a L’adaptation aux profondeurs.
- La théorie de l’adaptation chromatique complémentaire.
En surface, toutes les algues sont viables. Un peu en dessous (à partir de 10 mètres), les algues vertes meurent. Les radiations vertes restantes ne permettent que le développement des algues rouges. Cette théorie est sûrement vraie pour les zones pauvres en espèces.
-la théorie de la plasticité de l’appareil pigmentaire.
Pour cette théorie, il faut faire une étude du rendement synthétique.
Avant un certain point d’éclairement, la concentration en oxygène est inférieure à 0, il y a donc respiration.
Au dessus, la concentration en oxygène devient supérieure à 0, il y a donc plus d’O2 dégagé que d’O2 utilisé pour la respiration.
Si la lumière diminue, le point de compensation va diminuer.
Si le point de compensation diminue, cela permet à l’algue un développement dans une illumination plus faible.
En fait, seul le rapport de la chlorophylle A sur la chlorophylle B augmente. Il y a donc augmentation de la taille des antennes collectrices de photons.
Chaque espèce s’adapte différemment.
b Les contraintes liées à la vie en zone côtière.
Les marées vont provoquer une émersion des algues : il va y avoir une forte variation de l’illumination et une variation de pH et de salinité (on comprend alors le rôle de la paroi).
Les alginates sont des sucres acides avec des fonctions carboxyles qui piègent les ions en limitant l’entrée et la sortie d’ions au travers de la paroi.
L’illumination directe va entraîner la formation de radicaux libres (H2O2) qui entraînent une photo inhibition. Cette dernière est souvent irréversible.
Certaines algues vont développer un système de protection : présence d’un xanthophylle (violaxanthine) qui est transformé en zéaxanthine. La violaxanthine va modifier l’absorption car la zéaxanthine a un spectre d’absorption très fortement semblable à celui de la chlorophylle. Ce système est réversible quand la plante repasse à une plus basse illumination.
3 Zonation des algues.
Dans les zones de balancement de marées, il y a dispersion des algues selon les capacités de résistance aux contraintes liées à l’émersion.
Les phéophycées sont spécialisées dans l’adaptation.
Les Mycètes et les Lichens.
Généralités
Un plasmode est une masse cellulaire sans paroi.
Les thalles unicellulaires sont trouvés, par exemple, chez les levures.
Les thalles pluricellulaires peuvent être à filaments non cloisonnés ou à filaments septés (on trouvera un ou plusieurs noyaux selon l’organisme).
La multiplication végétative peut se faire selon diverses modalités.
On peut trouver une multiplication par des organes spécialisés comme avec les sporocystes qui libèrent des spores. Les sporocystes sont portés par des sporocystophores (filament érigé).Les spores obtenus peuvent être de deux types :
- Des spores endogènes qui donneront des spores sèches ou des spores flagellées si elles sont dans une cellule spécialisée.
- Des spores exogènes qui sont aussi appelées des conidies. Dans ce cas, le filament porteur est un conidiophore.
Les modes de reproduction sexuée existant sont divers :
- La planogamie.
Les gamétocystes (mâles ou femelles) donnent des gamètes flagellés qui sont émis à l’extérieur et qui vont fusionner. Cette fusion donne un planozygote qui se fixe.
- L’oogamie.
Le gamétocyste mâle (ou spermatocyste) donne des gamètes flagellés (spermatozoïdes) qui vont aller féconder le gamète femelle (oosphère) dans le sporocyste femelle (oocyste). L’œuf est contenu dans l’oocyste : c’est l’oospore.
- L’oogamie siphonogame.
Le gamétocyste mâle ne donne pas de spermatozoïdes. Il doit venir s’accoler au gamétocyste femelle puis émettre des siphons copulateurs qui perforent la paroi de l’oocyste.
- Cystogamie (ou zygogamie).
1 : progamétocystes + et - ;
2 : gamétocystes et suspenseurs ;
3 : fusion des gamétocystes ;
4 : zygospore.
Dans ce cas, on n’a besoin que de deux thalles compatibles, sans besoin de gamète. Il va y avoir émission d’un diverticule latéral (progamétocyste). Ensuite, apparaît une cloison latérale qui est le gamétocyste et le suspenseur.
Il va ensuite y avoir mélange des noyaux puis formation du zygote à paroi échinulée.
- La trichogamie.
Dans ce cas, le gamète mâle est une spermatie (non flagellé), émise par un filament.
L’organe femelle est un ascogone (cellule globuleuse surmontée d’un trichogyne). Il va y avoir fusion des parois puis injection du noyau mâle à l’intérieur de l’ascogone. Pendant le parcours du noyau mâle, ce dernier va se multiplier. Il y aura ensuite appariement sans fusion (dicaryon). On a alors des asques, aussi appelés, méiogamétocystes.
- La somatogamie.
Il va y avoir fusion de deux thalles compatibles et formations de dicaryons.
| |
Appareil végétatif
|
Multiplication végétative
|
Reproduction sexuée
|
|
Myxomycètes
|
Plasmode
|
|
|
|
Oomycètes
Chytridiomycètes
|
Filaments siphonnés
|
Zoospores
|
Oogamie,
Siphonogamie,
Planogamie.
|
|
Zygomycètes
|
Filaments siphonnés
|
Aplanospores
|
Zygogamie
|
|
Ascomycètes Filaments septés
|
Conidies
|
Trichogamie /Asques
|
|
Basidiomycètes Filaments septés
|
Conidies ou rien
|
Basides / Asques
|
I Phylogénie des champignons.
C’est à un règne à part comprenant environ 56 000 espèces connues (1 million d’espèces estimées). La classification est basée sur la morphologie, la biochimie et la biologie moléculaire.
A Les différents types de thalles.
Ils peuvent être macroscopiques souterrains (truffe) ou aériens (amanites). D’autres thalles sont microscopiques.
Tous les champignons sont des eucaryotes, se nourrissant par absorption ; leur appareil végétatif est un thalle. Ils ne sont pas chlorophylliens
On trouve les myxomycètes qui ont des mouvements amiboïdes car leur reproduction est réalisée par des sporocystes. On trouve aussi les oomycètes qui ont un thalle filamenteux, une nutrition par absorption et une reproduction par formation de spores.
B Le rôle des champignons dans la nature.
Les champignons peuvent être : décomposeurs, symbiotiques, parasites d’animaux ou de végétaux. Ils sont une source d’alimentation pour l’Homme, soit de manière directe soit indirectement (fermentation, affinage).
L’identification se fait selon des critères morphologiques : on doit donc connaître plus ou moins la classification.
C Les différents types de thalles.
Le thalle est la partie végétative assurant l’absorption des nutriments et permettant le développement des organes reproducteurs. Parmi les différents thalles, on distingue :
- Les plasmodes (myxomycètes)
- Les nématothalles : peuvent être érigés (allongés) ou coccoïdes.
- Les cladothalles : ils sont rares, comprennent les ascomycètes.
- Les thalles septés.
- Les thalles siphonnés : ont une cloison, pour délimiter un organe ou bine pour séparer une partie morte.
- Les agglomérats à hyphes : rigomorphes ou cordons (ressemblent à une racine) ; hyphes : cherchent les nutriments dans le sol, les parties anciennes s’agglomèrent pour former les hyphes.
- Les sclérotes : ce sont des amas d’hyphes dont des cellules de la périphérie se réunissent et les filaments centraux donnent la « moelle » riche en réserves nutritives. On les trouve chez les ascomycètes et chez les basidiomycètes. (Exemple : ergot du seigle).
- Les coussins conidifères : ce sont des agglomérats de conidiophores (Monilia laxa).
- Les acervules : ce sont les moins organisés. On les trouve sur les champignons de plantes. Les conidiophores s’accumulent sous l’épiderme du vaisseau (conidiophore).
- Les pycnides sont des structures en forme de bouteille avec un col qui forme un orifice.
- Les carpophores : ce sont des amas de filaments septés donnant un pied et un chapeau. Les hyphes sont serrés et forment un réseau lâche de filaments.
- Les stolons et rhizoïdes : ce sont des filaments qui poussent en ligne droite (stolon) puis qui se courbent. Il y a formation de rhizoïdes qui émettent une touffe de sporocystophores puis un nouveau stolon.
D Aspect cytologique du thalle.
Le noyau est petite taille, le nombre de chromosomes est restreint (entre 4 et  ; l’appareil de Golgi est atypique ; Présence de vacuoles dont l’importance augmente lors du vieillissement du thalle ; les réserves sont sous forme de lipides et de glycogène.
; l’appareil de Golgi est atypique ; Présence de vacuoles dont l’importance augmente lors du vieillissement du thalle ; les réserves sont sous forme de lipides et de glycogène.
E Composition de la paroi.
Elle est faite de 80% de polysaccharides, 10% de protéines et 10% de lipides.
La chitine, chez les Eumycota est un polymère de N-actéyl glucose amine (b1à4).
Le chitosan est formé de glucose amine sans groupement acétal.
Le glucane est un résidu de glucoses formant des polyoses liés en b1à3 et ramifié en b1à6.
F Croissance chez les champignons.
Il n’y a jamais de croissance intercalaire è Il y a toujours croissance apicale.
L’apex contient l’activité mitotique du filament et possède des propriétés biologiques particulières. Le diamètre d’un filament est constant. La paroi est rigide (indéformable) et il existe une polarité en direction de l’apex (mouvements de cytoplasme avec accroissement de la pression de turgescence).
G Organisation physiologique de l’apex.
1 : zone riche en vésicules ; 2 : une zone riche en mitochondries et REG et un certain nombre de filaments d’actine ; 3 : noyaux et vacuoles qui permettent d’apporter la pression de turgescence nécessaire à l’élongation.
Suivant les types de champignons, l’agglomération des vacuoles sera différente : diffuse, à l’extrémité. Les ascomycètes présentent une densité aux extrémités (= spitzenkörper). Chez les basidiomycètes, les spitzenkörper ne sont que des vésicules en anneau autour. On a une accumulation de vésicules impliquées dans la synthèse de la nouvelle paroi au niveau de l’apex.
Les vésicules migrent vers l’apex par des microfilaments d’actine. Ces vésicules fusionnent ensuite avec la membrane plasmique et il y a extrusion de leur contenu (polymères de la paroi).
Au niveau du dôme, il y a expulsion de monomères de chitine, ce qui crée une zone élastique. Sur le côté, il y a libération de la chitine-synthase qui va polymériser la chitine. Cette enzyme est expulsée au niveau du dôme.
H Formation des ramifications.
Si le diamètre est constant, le thalle ne peut pas grossir, il va donc se ramifier pour « évacuer » l’excédent de nutriments.
Après les premières cloisons, il y a formation des premières ramifications (indifféremment dans l’espace).
Il faut qu’il y ait « dissolution » de la paroi pour pouvoir former un nouvel apex : nécessité d’enzymes lytiques.
La ramification est aussi une alternative à la mobilité et à la dissémination. Ainsi, les colonies de champignons sont radiaires. Les parties vieilles sont au centre alors que les parties jeunes sont aux extrémités.
I Formation septums.
La formation des septums est centripète : elle commence au centre et s’étend vers les bords jusqu’à obturer les filaments. Les cloisons ont au milieu un pore permettant le passage du cytoplasme. Il sert à la communication des cellules et au passage des nutriments.
Quand il y a mort d’une cellule, il y a fermeture de ce pore afin d’isoler cette cellule.
II La multiplication végétative.
Un individu se développe identique à lui-même. On trouve :
- Les bourgeonnements : levures.
- Fragmentation du thalle
- Formation de cellules spécialisées.
A Bourgeonnement.
Il y a formation d’un bourgeon. Quand la cellule est aussi grosse que la cellule mère, elle peut se séparer ou bien rester accolée. Si elle ne se sépare pas, il y aura formation d’une file de cellules appelées pseudomycélium.
B Fragmentation du thalle.
Dans ce cas, les cellules formées sont des orthospores. Les cellules commencent par accumuler des réserves. Leur paroi se gélifie et les cellules se séparent du filament. Ces cellules deviennent des spores capables de générer un nouveau filament (géotricum).
Il y aura formation de clamidospores qui ont un rôle de conservation. Leur paroi est épaisse et apparaît double ; leur cytoplasme est riche en réserves et parait coloré.
C La formation de cellules spécialisées : sporulation vraie.
1 Les spores endogènes.
Ces spores apparaissent chez les oomycètes par un phénomène de polyplanétisme. La spore ne peut former directement un filament. La spore doit s’enkyster et donner une nouvelle spore flagellée (spore de second ordre). C’est le diplanétisme, avec deux spores nageuses.
Ces spores peuvent germer un nouveau filament ou donner, après un nouvel enkystement, d’autres spores (de troisième ordre). Ces ré enkystements successifs procurent la capacité de germer.
Les saprolégniales (inféodés à l’eau) voit leur phénomène de polyplanétisme s réduire au cours de l’évolution et de la diminution de leur dépendance à l’eau (aplanes).
2 Spores endogènes non flagellées.
Les spores non flagellées sont produites par les sporocystes, eux-mêmes, portés par des sporocystophores. On a trois types de sporocystophores :
- Type 1 : à columelle, contient de nombreuses spores.
- Type 2 : sans columelle, à nombre limité de spores.
- Type 3 : le sporocyste ne contient qu’une spore. La paroi du sporocyste se confond avec la paroi de la spore è sporocyste conidioïde.
3 Spores exogènes ou conidies.
Ces spores se forment directement sur les thalles (septés) : les spores sont inféodées au vent ou aux insectes.
Elles peuvent se former en chaîne et donner un filament court qui émet des conidies les unes à la suite des autres.
Ces spores peuvent aussi provenir de phialide. La phialide est une partie qui bourgeonne itérativement des conidies. La formation de conidies dans des phialides en goulot donne des endoconidies.
Les conidiophores enfermés dans un stroma sont des pycnides. Au fur et à mesure de leur formation, ils seront libérés. Ce fonctionnement ne concerne que les ascomycètes.
III La reproduction sexuée.
A Les générations morphologiques.
1 La multiplication végétative.
Le gamétophyte peut donner des gamètes haploïdes ou bien des spores équationnelles.
Le méiosporophyte donne des méiospores ou des spores équationnelles.
2 La reproduction sexuée.
Un cycle digénétique isomorphe présente deux générations identiques. Un cycle digénétique hétéromorphe présentera deux générations distinctes morphologiquement.
a Cycle monogénétique haplophasique.
On observe un gamétophyte haploïde.
Les gamètes vont fusionner pour donner un embryon qui subit immédiatement la méiose et donne quatre cellules (spores) qui pourront subir des phases de multiplication végétative.
b Cycle monogénétique diplophasique.
Il y a formation d’un embryon par une fécondation. Celui-ci pourra subir deux phases de reproduction sexuée. Des cellules spécialisées vont subir la réduction chromatique et immédiatement fusionner (fécondation).
On observe un méiosporophyte diploïde.
c Cycle trigénétique haplodiplophasique.
La réduction chromatique donne des spores qui restent à l’état haploïde sans fusionner. Elles peuvent rester en l’état durant un long moment. Quand elles fusionnent, elles donnent une espèce diploïde. Dans la seconde partie du cycle, on peut observer des étapes de multiplication végétative. La troisième phase comprend la fin de l’individu diploïde qui subit la réduction chromatique.
Suivant le moment de la méiose et de la fécondation, on distinguera des phases nucléaires.
3 Exemples de cycles.
La fécondation, une planogamie, donne un planozygote qui va se poser, puis germe. En germant, il y a formation d’un nématothalle qui libère directement des zoospores diploïdes. Indirectement, il peut y avoir libération de zoospores haploïdes par l’intermédiaire d’un sporokyste (avec réduction chromatique).
Le cycle est isomorphe.
b Saprolegnia monica.
La reproduction sexuée se fait par des sporocystes qui libèrent des zoospores flagellés (polyplanétisme). La fécondation est une oogamie siphonogame.
Le thalle, diploïde, va former des organes de reproduction : les oocystes chez les femelles (contiennent des oosphères) ou des spermatocystes chez les mâles (fécondation par les noyaux).
La fécondation donne des zoospores à 2N qui vont germer et donner un thalle diploïde.
c Plasmopara viticola.
Dans le gamétophyte femelle, on ne trouve que des noyaux. Un seul œuf est formé.
d Mucor mucedo.
Les spores haploïdes, servant à la multiplication asexuée, sont disséminées par le vent.
Quand il y a rencontre de deux gamétophytes compatibles, deux gamétocystes vont fusionner (caryogamie). On obtiendra alors, dans la zygospore, un seul noyau, diploïde, suspendu entre un suspenseur et le gamétocyste.
La zygospore, par réduction chromatique, donne deux spores + et deux – (haploïdes).
e Les Ascomycètes, Peziza.
Le thalle est haploïde, septé : c’est un mycélium primaire.
L’organe femelle est un ascogone (une cellule renflée, terminée par un poil, le trichogyne). Les organes mâles sont les anthéridies (réalisent la fécondation par les noyaux).
Une fois l’ascogone atteint, on ne voit pas de fusion des noyaux : il y a mise en place du syncarion.
Il y a formation de dangeardies par le phénomène du crochet. L’asque est le méiosporocyste.
Le cycle est digénétique haplodiplophasique. Le gamétophyte débute le cycle.
Pendant la formation des asques, il y a migration des noyaux vers le haut. A la base, il y a mise en place de vacuoles qui poussent les noyaux : il y a formation d’une vésicule ascale à la périphérie du plasmalemme.
Après la mitose, cette vésicule (ascale), se fractionne pour donner la membrane plasmique de chaque future spore. Les ascospores sont endogènes.
La libération de ces spores va se faire par éclatement de la structure de l’asque. La paroi est formée de deux tuniques : la tunique externe est l’exoascus (tunique hydrophobe, rigide), la tunique interne est l’endoascus (tunique mince, lipophile, souple).
On trouvera des asques bituniqués (ils ont deux tuniques visibles après coloration) ou des asques unituniqués (les tuniques sont collées et l’on n’en voit qu’une).
Les différents mécanismes d’ouverture des asques sont :
- Jack in the box : Les vacuoles poussent et font craquer la tunique externe. La tunique interne va sortir. Ce moyen d’ouverture est typique des asques bituniqués.
- Eclatement.
- Ouverture par anneau.
- Ouverture par clapet.
f Les hémibasidiomycètes : Puccinia graminis.
Puccinia donne la rouille du blé.
Le cycle est trigénétique.
Le cycle commence avec un gamétophyte qui va infester l’épine vinette. On trouve un épithélium haploïde sur la face supérieure (gamétophytes mâles) ainsi que sur la face inférieure (gamétophytes femelles).
Les spermaties (non flagellées, immobiles) sont libérées avec une goutte de miellat pour attirer les insectes. Il y a mise en place d’hyphes flexueux pour récupérer les spermaties (d’origine différente).
Ces hyphes flexueux sont en contact avec des écidies (appareil femelle). Les spermaties se collent à l’hyphe. Le noyau mâle entre et descend vers le noyau femelle. Il va y avoir formation d’un dicaryon avec mitoses successives et bourgeonnement (les écidiospores).
Ces écidiospores vont infecter un deuxième hôte (le blé) et former un mycélium dicaryontique. Au bout d’un moment, il va y avoir différenciation des urédosores qui vont émettre des urédospores. Ces urédospores sont des cellules rondes à paroi épaisse, avec deux noyaux, et reliées à la feuille par un pédicelle. Les urédosores sont des zones de regroupement des urédospores.
A la fin de l’été, le mycélium forme un nouveau type de spores : les téleutosores (taches noires) où l’on trouve des téleutospores. Ces spores sont pédicellées avec deux noyaux qui subissent la caryogamie.
Au printemps, il y a émission d’un filament (boyau) où un noyau diploïde passe à l’intérieur. Il y a dans le boyau, formation de quatre gamètes haploïdes (méiosporocystes = basides). Chaque noyau est isolé par une cloison transversale.
Chaque cellule à noyau haploïde bourgeonne une petite cellule où vient le noyau : les basidiospores. Ces dernières formes sont légères et vont réinfecter l’épine vinette.
La première génération est un mycélium haploïde.
La seconde génération est composée des écidiospores et des urédospores.
La troisième génération comprend les téleutospores et les basides (ou méiosporocystes).
g Les basidiomycètes, Coprinus comatus.
La fécondation est une somatogamie.
Le mycélium primaire (haploïde) va faire une fécondation sans différenciation d’organes sexuels : il y a fusion de thalles complémentaires (à plasmogamie). Il y a formation d’une cellule à dicaryon (pas de fusion). Il vient ensuite une anse d’anastomose (»dangeardie). Il va y avoir une succession de cellules à dicaryon : le mycélium reste marqué par la bouche mycélienne, c’est le filament secondaire. C’est ce filament secondaire qui va former le carpophore.
Au pied, il reste une membrane venant de la collerette qui se déchire : la volve. Au-dessus de cette volve, on trouve une cicatrice (un anneau).
Les lamelles sont des filaments dicaryontiques. Ce sont aussi des zones fertiles. Elles constituent l’hyménium :
- Cystites (réserves)
- Cellules sécrétrices (donnent du latex)
- Basides (cellules où a lieu la caryogamie).
Dans les basides a lieu la méiose et donne quatre cellules à N qui donneront quatre pointes (les stérigmates) où vient migrer un noyau. C’est une basidiospore qui va germer un mycélium haploïde. Sur les quatre basidiospores on trouve deux cellules + et deux cellules -.
Chez les Auriculariales, le thalle est cloisonné. On a les basides en position latérale puis émission d’un long boyau d’où sortent les basidiospores.
Les Urédinales et les Auriculariales ont des archéobasides. Les Tremellales ont des hétérobasides. Les Agaricales ont des basides typiques.
è Les champignons montrent des formes très originales :
- On trouve des caryogamies différées.
- Il y a absence de gamètes mâles. Ils sont remplacés par des conidies (spores de multiplication végétative).
- Il peut y avoir autogamie, avec deux gamètes femelles.
- La somatogamie est une fusion de deux thalles qui donne un mycélium à dicaryon.
B Homothallisme et hétérothallisme.
A Hétérotrophie vis à vis de l’azote.
Parmi les autotrophes, on peut trouver des levures, qui, dans certaines conditions, vont pouvoir utiliser l’azote atmosphérique. Des mycètes comme Aspergillus niger et Mucor mucedo vont absorber de l’azote nitrique
D’autres individus sont appelés semi-hétérotrophes (Mucor sp., des levures) et sont caractérisés par une consommation d’azote ammoniacal.
Les hétérotrophes peuvent être des saprophytes comme Saprolegnia, des symbiotiques comme les champignons lichénisants, des parasites comme Phytophtora, Plasmopara ou bien des prédateurs comme Dactylella acrochaete. Ces individus consomment de l’azote organique.
B L’hétérotrophie vis à vis du carbone.
1 Le saprophytisme.
On peut trouver trois types de pourriture :
- Pourriture cubique.
- Pourriture fibreuse.
- Pourriture alvéolaire.
2 Le parasitisme.
Comme type de parasitisme, on peut citer la graphiose de l’orme (maladie de l’orme champêtre), la maladie de la pomme de terre irlandaise.
Ce parasitisme peut s’effectuer par une effraction tissulaire et même parfois, cellulaire (avec la formation de suçoirs). Elle peut aussi se présenter par une action biochimique avec la sécrétion d’enzymes, de toxines, de substance de croissance. Ces actions provoquent un flétrissement (les toxines) ou un gigantisme (substances de croissance).
On trouve champignons parasites de faiblesse.
Ces champignons se développent sur une plante même quand elle est morte : ce sont des nécrotrophes ou bien saprophytes. Il existe différents types de biotrophie avec aussi des développements sur des plantes vivantes. Ce sont des parasites forts ou obligatoires.
Les différentes étapes nécessaires à une infection :
- Reconnaissance de la plante par échange de signaux biochimiques. On trouve des spectres étroits ou bien plus larges. (spectre étroit pour les parasites forts et large pour les parasites faibles).
- Attachement à la surface de la plante.
- Effraction tissulaire.
- Développement interne, avec effraction cellulaire possible.
- Troubles chez la plante.
- Reproduction du champignon.
a L’adhésion.
Cette adhésion se fait par sécrétion d’un mucilage de polysaccharides et de glycoprotéines sous le tube germinatif. Il va y avoir synthèse d’enzymes pour dégrader l’épiderme de la plante.
b La pénétration.
La pénétration peut se faire par voies naturelles (comme les stomates) et dans ce cas, il n’y a pas de différenciation d’un tube germinatif, au niveau de l’épiderme ou des racines, ce qui est une pénétration directe. Dans ce dernier cas, il va y avoir mise en place d’organes particuliers (actions mécaniques) et libération d’enzymes pour dégrader la paroi (actions chimiques).
Pour l’action mécanique, il va y avoir renflement du tube germinatif (ou appressorium) : la pression osmotique du champignon est plus forte que celle de la plante afin d’y résister.
Il va ensuite y avoir formation d’un filament fin (point d’infection) duquel partent des filaments infectieux qui portent les suçoirs (Haustorium).
Ce sont les structures infectieuses.
Les champignons à appressorium mélanisés vont renter sans problème. Les formes à appressorium muté ne rentrent pas. La mélanine permet l’accroissement de la pression pour la pénétration.
Lors de l’action chimique, la formation du tube germinatif entraîne la formation d’un mucilage, d’enzymes hydrolithiques pour la dégradation de l’épiderme de la plante (cutinases, lipases, cellulases…) Il va y avoir une diminution de la paroi du végétal.
La pénétration mécanique par les racines.
Deux cas existent pour cette pénétration : coussins d’infection et rhizomorphes.
Les coussins d’infections sont des agglomérations d’hyphes d’où partent plusieurs points d’infection (les coins d’infections). Ils partent en parallèle et attaquent la plante.
Les rhizomorphes sont des extensions gélatineuses qui permettent l’adhérence. Ce sont des émissions latérales qui pénètrent car ce sont des filaments agglomérés.
Une fois à l’intérieur, deux cas sont possibles :
- Nécrotrophe : il y a dégradation des parois du végétal puis des parois cellulaires. Les filaments s’insinuent dans toutes les fentes mais provoquent aussi la mort de la plante.
- Biotrophe : Les filaments migrent dans la plante, au niveau de la lamelle moyenne. Seuls les suçoirs entrent dans les cellules. Ces derniers permettent aussi la fixation.
Chez Oïdium, champignon externe, les suçoirs forment des structures d’adhésions.
Les toxines vont provoquer un flétrissement de la plante. Les substances de croissance, provoquent elles, un gigantisme mais aussi un nanisme et une castration sont possibles.
3 L’endophytisme.
L’endophytisme est entre le parasitisme et la symbiose. C’est un cas qui provoque peu de dégâts.
a Chez Claviceps.
L’infestation ne provoque aucun symptôme, sauf à la floraison où dans l’ovaire, le stroma donne des conidies. Le stroma va se différencier en sclérote qui tombe au sol. Là, il se développe des structures de reproduction sexuée.
b Chez Epichlöe triphina (la quenouille de graminées).
Il va se former un stroma blanc autour de la tige. C’est là où sont les structures de reproduction.
c Chez Sphacelia tryphina.
Ce champignon est responsable de l’ivraie enivrante. Il se développe des graines qui donneront une plantule avec le champignon. Les nouvelles graines porteront toujours le champignon.
Les animaux qui consomment ces graminées auront des malaises et autres symptômes similaires car ces champignons synthétisent des alcaloïdes.
4 Les symbioses.
On distinguera deux cas de symbioses : la symbiose par les mycorhizes ou par les lichens.
Les mycorhizes se trouvent chez les ascomycètes, les basidiomycètes ou chez les zygomycètes (Glomales, la reproduction sexuée est diminuée).
Dans le cas de l’ectomycorhize, le champignon se développe à l’extérieur ; il ne se trouve jamais dans les cellules ; il provoque des modifications morphologiques des racines ; il se développe autour des racines.
Dans le cas de l’endomycorhize, le champignon ne montre pas de développement important à l’extérieur ; il entre à l’intérieur des cellules ; il n’entraîne pas de modifications importantes des racines.
a L’endomycorhize (Les Glomales).
Les glomales sont uniquement symbiotiques. On trouve quatre familles : Glomus, Gigaspora, Sclérocystis, Acannospora.
Une spore va germer et donner, à l’extérieur, un filament et un appressorium (comme chez les parasites). Il se forme des arbuscules dans les cellules végétales (» suçoirs). Dans certaines familles comme Glomus, il y a formation de grosses vésicules contenant des réserves pour le champignon : les méats.
Les arbuscules sot des surfaces d’échange entre le champignon et la plante (Gigaspora).
La plante va former une matrice extra-fibrillaire pour empêcher la prolifération des arbuscules (durée de vie : 3-4 jours).
Les mycorhizes d’Ericaceae.
La racine est fine à petit diamètre et simple (cortex, endoderme, pôle ligneux). La colonisation du champignon ne se fait que dans les cellules corticales. Les racines mycorhizées vont dégénérer au fur et à mesure.
b L’ectomycorhize.
Cette forme de symbiose fait intervenir les basidiomycètes et les ascomycètes (Tuber melanosporum).
Il va y avoir formation d’un réseau extra-matriciel (mycélium dans le sol) et d’un réseau interne (réseau de Hartig). Il n’y a pas de pénétration intracellulaire (reste dans les méats).
La racine va se transformer.
Le réseau de Hartig est une surface d’échange entre le champignon et la plante.
c L’ectendomycorhize.
Les mycorhizes à Orchidée.
Un basidiomycète dont la forme végétative est Rhizoctonia va coloniser les racines adultes et l’embryon (dépourvu de réserves). La colonisation de l’embryon permet sa germination. Il faut un équilibre entre la phagocytose et le parasitisme pour que l’embryon puisse atteindre la germination.
è C’est un état de parasitisme contrôlé pour une vie commune.
Cette ectendomycorhize est un type intermédiaire.
Il y a formation d’un manchon et une pénétration intracellulaire.
d Conclusion sur les symbioses.
La symbiose permet aux champignons d’obtenir des synthétats, source de carbone. La plante, grâce au champignon, peut aller chercher l’eau et les sels minéraux (P, Cu, Z, S) beaucoup plus loin.
Certains champignons vont même jusqu’à récupérer des ions comme NO3- et bien d’autres pour que la plante puisse se développer sur de mauvais milieux.
Il y a aussi l’action des hormones de croissance.
Les Herbacées sont endomycorhizables alors que les Arbres sont ectomycorhizables.
5 La place des champignons dans la biosphère.
Les saprophytes permettent la dégradation des plantes mortes et vont former l’humus et donc, améliorer les sols.
L’endomycorhize permet l’accroissement de l’absorption de l’eau et des sels minéraux.
Les cellules lignivores participent à la nutrition des herbivores.
Les parasites participent à l’équilibre des populations végétales.
V La symbiose lichénique.
Un lichen est une association stable entre une algue et un champignon qui donne au final, un nouvel individu.
Les lichens sont des organismes polyphylétiques. Ils sont apparus à 5 reprises dans l’évolution. Les plus vieilles formes ont environ 400 millions d’années (au Dévonien). On en connaît 25 000 espèces.
Le partenaire fungique est appelé « mycobionte ». Ce sont essentiellement des ascomycètes (et souvent des Pyrénomycètes et des Discomycètes). On trouve tout de même 1% de basidiomycètes. 13250 espèces de champignons sont lichénisantes.
Chez les algues, photobiontes, on trouve 40 espèces lichénisantes. Exemple : Trebouxia, Pseudotrebouxia, Trentepholia, Nostoc.
A Les diverses formes de lichens.
1 Les lichens gélatineux.
Ces lichens sont homomères. Les hyphes sont mélangés aux cellules des algues. Ils sont homogènes dans le désordre (en général, Ascomycètes + Nostoc).
2 Les lichens secs.
Ces lichens sont toujours organisés en strates. Ils sont appelés hétéromères (structure hétéromère stratifiée sauf chez les lichens fructiculeux). Parmi eux, on trouve :
- Les lichens secs crustacés : Ces lichens sont sur les rochers (rupicoles), les trottoirs. Ils sont complètement dans leur substrat. De l’extérieur (air) vers l’intérieur (contre le substrat), on trouve : un mycélium (le cortex), une couche gonidiale et une médulla arachnoïde.
- Les lichens secs foliacés : Ces lichens sont sur toute la longueur du substrat. De l’extérieur vers l’intérieur, on trouve : un cortex supérieur, une couche gonidiale (cellules algales), une médulla arachnoïde et un cortex inférieur avec des rhizines.
- Les lichens secs fructiculeux : Ces lichen fructiculeux pendent des branches d’arbres. Ils ne sont appliqués au substrat que par un point. Ils ont, en coupe, une forme relativement cylindrique. L’extérieur vers le centre, on trouve : un cortex supérieur, une couche gonidiale (avec cellules algales), une médulla arachnoïde puis une chondroïde. Ces lichens ont une structure radiée.
3 Le thalle secondaire.
Ce thalle secondaire est appelé podétion.
En coupe, un podétion présente : un cortex supérieur, une couche gonidiale, une médulla arachnoïde creuse, une médulla chondroïde et une lacune centrale.
Les cyphelles sont des trous pour l’aération de la couche gonidiale.
Les céphalodies apparaissent quand deux lichens sont superposés. Un champignon, une chlorophycée et une cyanobactérie. On a donc un thalle à cyanobactérie et un thalle à chlorophycée ainsi qu’un double champignon. Il peut y avoir fixation d’azote atmosphérique grâce aux cyanobactéries.
B Les structures de multiplication et reproduction.
Les pycnides. Ce sont des structures de multiplication du champignon qui forment et libèrent des conidies.
Les isidies. Ce sont des structures dressées, ramifiées à la surface du thalle. Elles forment de petits cylindres constitués d’algues, entourés de filaments mycéliens. Elles sont détachables au vent.
Les sorédies. Les sorédies sont des cellules d’algues individuelles, entourée d’hyphes regroupés dans une soratie.
Le champignon, seul, possède des structures d reproduction :
- Des apothécies si ce champignon est un discomycète. Elles sont sur les bords du thalle et forment un petit sillon convexe ou concave.
- Des périthèces si le champignon est un pyrénomycète. Elles prennent la forme d’une urne, seule, ou en groupe (se trouvent surtout chez les lichens crustacés).
Quand on a un asque, une ascospore doit rencontrer une algue, sinon, elle meurt. Si elle tombe sur une surface humide et que le thalle est blessé, il y aura symbiose dans le cas d’une rencontre avec une algue compatible.
Croissance et longévité.
Les crustacés ont une croissance de 2 à 3 millimètres par an. Les autres lichens ont une croissance comprise entre 3 et 4 centimètres par an.
La croissance est composée de plusieurs phases :
- Phase I : croissance optimale.
- Phase II : phase de maturation.
- Phase III : phase de dégénérescence (au centre) alors que l’on observe une croissance de l’extérieur.
Si le lichen est affaibli (par la pollution par exemple), il pourra y avoir installation d’un parasite.
Contre ces parasites, les lichens vont former des zones encerclantes et synthétiser des antibiotiques et des substances métaboliques secondaires (les acides lichéniques).
C La symbiose lichénique.
1 Symbiose contre parasitisme.
Le champignon absorbe les photosynthétats de l’algue. Celui-ci amène l’eau, protège de la dessiccation, des UV et du broutage par les herbivores grâce aux acides lichéniques.
2 Le parasitisme.
Dans ce cas, c’est le champignon qui va parasiter l’algue : on parle d’hélotisme. L’algue est prisonnière dans le champignon. Il y a inhibition de sa reproduction sexuée et sa photosynthèse est modifiée en quantité et en qualité.
3 L’algue parasite le champignon.
Les céphalodies sont considérées, par certains, comme des galles. On observe des réactions de défense du champignon contre l’algue.
è En fait, on observe un état d’équilibre et des relations mutualistes.
L’antagonisme du champignon est compensé.
è Il y a synthèse d’un nouvel individu : le LICHEN.
La somme du champignon et de l’algue fait apparaître des propriétés propres aux lichens.
4 Relation morphologique entre algue et champignon.
On distingue trois types :
- Cohabitation sans contact entre les cellules de l’algue et celles du champignon.
- Le champignon enserre l’algue. Pour cela, il va former des crampons.
- Haustorium. Si l’algue est une cyanobactérie, le champignon peut entrer dans le mucilage, mais pas dans les cellules. Si l’algue est une chlorophycée, le champignon peut faire un Haustorium qui entrera dans la chlorophycée.
5 Relations biochimiques.
Les algues fournissent les photosynthétats (1/3 dans le cortex et 2/3 dans la médulla).
Quand le champignon récupère les sucres, il va les transformer en sucre-alcool (ribitol, mannitol…). Ces sucres modifiés permettent le maintien d la turgescence du lichen et le protège d’une dessiccation trop élevée.
Les champignons vont synthétiser des métabolites secondaires (antibiotiques et acides lichéniques). Ces acides vont intervenir dans la régulation du nombre de cellules d’algue (en le faisant diminuer). Toutefois, ils protègent aussi les algues contre les UV et contre la dessiccation. Les champignons interviennent aussi dans la protection contre les métaux lourds.
La classification est basée sur la classification des champignons, puis sur les modes de reproduction…
D Les lichens dans leur milieu.
Les lichens ont une teneur en eau très faible (2 à 10% du poids sec du lichen). S’ils se dessèchent, il y a diminution de la photosynthèse. Le cortex va alors s’épaissir, devenir opaque et l’algue ne reçoit plus d’UV. Ils peuvent toutefois se réhydrater très vite : poïkilohydre. Il y aura alors reprises des fonctions métaboliques. Le cytoplasme doit être résistant.
Il n’y a photosynthèse que durant les premières heures d’ensoleillement. Dès que le climat devient trop sec, il y a arrêt de la photosynthèse. Ce qui explique la lente croissance des lichens.
Les lichens sont quand même capables de coloniser des milieux extrêmes : on les trouve dans la toundra, dans l’antarctique (350 espèces de lichens contre deux espèces deux plantes vasculaires). è Ce sont des pionniers.
Les facteurs écologiques pour l’installation des lichens :
- Types de substrats (milieu, composition…)
- Climats (humidité, ensoleillement)
- Facteurs biologiques : animaux intervenant sur leur installation (déjection) et pollution humaine.
1 Les lichens terricoles.
Ces lichens sont les plus soumis à des modifications de substrat. Ils sont sur l’humus, le calcaire, avec des végétaux, avec des débris organiques.
Si l’on se trouve dans des prairies semi-désertiques, on a une espèce de Pormélia (foliacée) qui va donner une espèce de Cladonia puis disparaître. Cladonia (Podétiée) donne à son tour 4 à 5 espèces nouvelles de Cladonia mais l’espèce d’origine disparaît. Sur ces 4 ou 5 espèces, 3 ou 4 vont disparaître et une survivra.
2 Les lichens épiphytes.
On trouve des successions lichéniques différentes si le végétal est isolé ou en groupe (forêt). L’espèce du végétal colonisé intervient aussi. Les écorces lisses à pH alcalin hébergeront plutôt des lichens foliacés alors que les écorces rugueuses, à pH acide, hébergeront des lichens fructiculeux.
3 Les lichens lithicoles.
On trouve différents lichens selon la nature chimique. A pH basique, on aura un lichen comme Collema alors qu’à pH acide on trouvera des crustacés.
On distinguera aussi des lichens calcicoles et des lichens calcifuges.
La nature chimique pourra être modifiée par l’humus, les déjections d’oiseaux (chargées en azote).
La nature physique pourra être modifiée par les caractères des failles.
4 Les lichens aquatiques.
Ils sont sur les roches acides et en milieu marin. Ils sont différents en fonction des zones de balancement des marées.
Au niveau de la zone intertidale, on trouve Verrucaria qui forme une ligne noire.
Lichina forme une ligne brune avec les embruns, les marées, les équinoxes.
Calauplaca et Xanthoria forment une ligne orange.
b Les lichens d’eau douce.
Il existe des lichens émergés 6 mois par an, d’autres émergés seulement 3 mois par an et d’autres qui ne sont jamais émergés.
Les lichens n’ont pas le même système de filtration : pour cela, il y a utilisation des acides lichéniques.
Les pollutions par H2SO4 (ou d’autres polluants) peuvent tuer les lichens.