Sommaire du cours de Physiologie Végétale.
|
|
|
|
| Définition |
|
|
Les différents végétaux
|
Partie 1 : Nutrition et métabolisme
|
|
Particularités des végétaux
|
|
|
|
1. Au niveau de l’individu
|
1. Anabolisme et catabolisme
|
Partie 2 : Développement
|
|
2. Au niveau de la cellule
|
2. Autotrophie et hétérotrophie
|
|
|
3. Au niveau de l’organisation des tissus et organes
|
3. Le couplage énergétique
|
A. Sites et formes de croissance |
| |
4. Les composés riches en énergie
|
B. Cinétique de croissance |
| |
|
C. Variation dans la croissance |
| |
A. Méthodes de détermination |
|
| |
B. Résultats |
1. Les phytohormones |
| |
|
2. Le phytochrome |
| |
Introduction |
|
| |
1. La photosynthèse : assimilation chlorophyllienne |
A. Tropismes |
| |
2. La photosynthèse Au niveau de la plante |
B. Tactismes |
| |
3. L’hétérotrophie par rapport au carbone |
C. Nasties |
| |
Conclusion |
D. Nutations |
| |
|
E. Autres mouvements |
| |
A. Les différentes formes d’azote présente dans le végétal |
|
| |
B. Les différentes formes d’azote disponible dans la biosphère |
1. Cycle de développement des spermaphytes |
| |
C. Assimilation de l’azote minéral par la plante |
2. Développement embryonnaire : la germination |
| |
D. Utilisation de l’azote atmosphérique |
3. Différenciation des autres organes |
| |
E. Le cycle de l’azote |
4. Capacités organogènes des végétaux |
| |
|
5. Développement reproductif |
| |
A. L’eau du sol |
|
| |
B. L’eau dans la plante |
|
| |
|
|
| |
A. Les ions minéraux : nature et importance |
|
| |
B. Origine des ions minéraux |
|
| |
C. Modalités et mécanismes de l’absorption |
|
| |
D. Rôle des ions minéraux dans la plante |
|
INTRODUCTION GENERALE
I Définition.
La physiologie végétale est l’étude des mécanismes qui régissent le fonctionnement et le développement des végétaux. On a deux grandes parties :
- Nutrition et métabolisme : * c’est l’acquisition des éléments indispensables à la vie, * la transformation de ces éléments par leur intégration dans la matière organique (dans la biomasse).
- Mécanismes pour le passage de la graine (état de vie ralentie) à l’état reproducteur (cycle du végétal : croissance et développement).
II Les différents végétaux.
Les procaryotes sont à l’origine des halobactéries (vivent en milieux salins ou saumâtres). Ce sont les premiers organismes à utiliser l’énergie lumineuse. Avant, les cellules étaient toutes hétérotrophes. Ces halobactéries possèdent un pigment spécial : le Rétinal, qui, associé à une protéine, forme la bactériodiopsine (pigment comparable à ceux de l’œil). Ce pigment absorbe la lumière et libère des protons qui se répartissent de part et d’autre de la membrane en formant un gradient électrochimique (ce gradient va de l’extérieur vers l’intérieur). Grâce au couplage avec une ATPase, il y a formation d’énergie (ATP) disponible pour les synthèses.
 C’est le premier système de production d’énergie chimique photodépendante. Ce processus est toutefois différent de la photosynthèse.
C’est le premier système de production d’énergie chimique photodépendante. Ce processus est toutefois différent de la photosynthèse.
La photosynthèse : c’est une utilisation de l’énergie lumineuse pour pouvoir produire de l’énergie chimique (ATP) et du pouvoir réducteur (NADPH,H+ ou NADH,H+) en vue de réduire le CO2 atmosphérique.
Les bactéries photosynthétiques possèdent différents pigments :
- Les photoautotrophes ou photolithotrophes sont composés de deux groupes :les thiobactéries de couleur pourpre et les chlorobactéries de couleur verte.
- Les photoorganotrophes sont des athiobactéries de couleur pourpre qui utilisent l’énergie lumineuse et la matière organique.
L’énergie solaire absorbée, excite des électrons qui sont transmis le long d’une chaîne rédox inverse. Au cours de ce flux d’électrons, le gradient électrochimique de protons sert à produire de l’ATP (énergie chimique) et du pouvoir réducteur (équivalents réduits) nécessaire pour réduire le CO2 atmosphérique. Les équivalents réduits sont des composés qui cèdent des protons et/ou des électrons.
Chez les bactéries, les éléments qui permettent la réduction sont :
- Des composés minéraux comme H2S où H+ sert à la réduction
- Des composés organiques, AH chez les photoorganotrophes (athiobactéries).
Chez les bactéries actuelles, la production d’équivalents réduits nécessite le transfert d’électrons avec consommation d’ATP.
Au cours de l’évolution, l’atmosphère s’est enrichie en oxygène grâce à la photosynthèse. Les bactéries actuelles ne représentent pas les systèmes primitifs. L’atmosphère est devenue oxydante grâce à l’oxygène : la sélection a fait apparaître un système qui utilisait l’oxygène : la chaîne respiratoire.
Remarque : les bactéries ont un seul photosystème alors que les algues-bleues et las plantes supérieures en ont deux.
Tous ces phénomènes peuvent se ramener à des systèmes rédox : importance de la notion d’énergie. Chez les végétaux supérieurs, ont a deux organites importants : le chloroplaste et la mitochondrie. Ces deux plastes ont une origine endosymbiotique. La mitochondrie viendrait de l’association d’une bactérie et d’une cellule à fonctions respiratoires alors que le chloroplaste aurait pour origine une algue bleue ou un organisme proche.
Comme preuve de cette origine symbiotique, on voit que ces deux organites ont une double membrane, de l’ADN et leurs divisions sont autonomes. De plus, ces deux organites sont sensibles aux antibiotiques.
Un végétal chlorophyllien est un organisme chlorophyllien autotrophe qui pourra fabriquer les constituants fonctionnels et structuraux dont il a besoin à partir de molécules inorganiques.
III Particularités des végétaux.
A Niveau individuel.
Les végétaux sont immobilisés, ce qui les obligent à s’adapter au milieu, à la disponibilité en eau, à la lumière et à la température et à se nourrir de ce qui les entourent (CO2 gazeux, eau liquide, éléments minéraux). Ce sont particularités qui définissent l’autotrophie.
B Niveau cellulaire.
Toutes les cellules eucaryotes vont être sensiblement identiques. Cependant, chez les végétaux supérieurs, une cellule adulte se distingue par la présence :
- La présence d’une paroi squelettique rigide mais vivante (composée de polysaccharides, de protéines, d’enzymes) qui est un squelette péricellulaire impliqué dans de nombreuses fonctions d’échanges (de cellule à cellule ou avec le milieu), de reconnaissance, et de défense.
- La présence d’une grande vacuole (résultant de la fusion de plusieurs petites) qui est un réservoir d’eau indispensable à la turgescence de la cellule et au port de la plante. C’est aussi un lieu de stockage de composés minéraux ou organiques qu’ils soient des réserves ou des déchets. Cette vacuole permet la détoxication de la cellule par la séquestration de composés toxiques en son intérieur.
- La présence de plastes (surtout dans les parties aériennes) qui permet la conversion de l’énergie lumineuse en énergie chimique. Ces plastes proviennent de l’évolution de protoplastes
- Les cellules sont théoriquement toutes totipotentes car elles contiennent la totalité de l’information génétique qui leur permet de se différencier en un nouvel individu. Cette capacité est utilisée dans les cultures in-vitro et dans les biotechnologies. Pour réaliser des cultures de végétaux à partir de cellules isolées, on se sert généralement de protoplastes car ce sont des cellules sans paroi.
- L’information génétique se trouve dans le noyau, les mitochondries, les chloroplastes. Les inhibiteurs agissent sur toute la cellule (ou les cellules) alors que les antibiotiques n’agissent que sur l’ADN chloroplastique et mitochondrial.
C Au niveau de l'organisation des tissus et des organes.
Partie 1 :
NUTRITION ET METABOLISME.
RAPPELS
Le métabolisme (transformation de la matière organique) est l’association du catabolisme (phase de dégradation) et de l’anabolisme (phase de synthèse).
La particularité des végétaux supérieurs est leur catabolisme (production de matière première) qui a un rendement énergétique réduit. Ce faible rendement est compensé par l’énergie solaire. Les lipides, glucides et protides ont des propriétés énergétiques différentes selon la lumière et la quantité d’oxygène. Un gramme de protide ou de lipide libère 4 KCal, alors qu’un gramme de lipide libère neuf KCal.
Thermodynamique :
• Enthalpie constante : 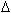 H
H
• Entropie plus ou moins ordonnée dans un système : S G=H-TS (mesure de la variation d’énergie). Si la pression et le volume sont constants, on a l’énergie libre qui est égale à G.
• Enthalpie et potentiel électrochimique :
G’°=-RT.log(K) E’°=RT/nF.log(K) G’°=-nF.E’°
Couplage énergétique : Les réactions de l’anabolisme et les premières étapes du catabolisme sont endoénergétiques. Elles doivent recevoir l’énergie d’autres réactions. G d’un système doit toujours être inférieur à zéro. Chez les végétaux supérieurs , cette énergie vient en partie de l’énergie lumineuse. K=(C.D)/(A.D) ; G=G’°+RTlog(K). Souvent les réactions sont des réactions d’oxydoréduction où l’ion majeur est H+.
Partie 1 :
NUTRITION ET METABOLISME.
Chapitre 1 : DETERMINATI0N DES BESOINS ALIMENTAIRES DES VEGETAUX
RECHERCHE DES ELEMENTS ESSENTIELS
A Méthode de détermination.
1 Méthode analytique : analyse élémentaire.
Cette analyse donne des renseignements sur la composition globale des éléments absorbés par une plante, ce qui ne signifie pas que cet élément soit important ou indispensable. Certains éléments du sol seront absorbés passivement par la plante. Cette méthode doit être accompagnée par une méthode d’étude synthétique.
2 Méthode synthétique.
On réalise une solution synthétique afin d’identifier la solution la plus simple qui pourra conduire au développement normal du végétal (c’est une solution nutritive). On obtient ensuite un classement en trois groupes des éléments disponibles dans le sol : essentiels, indifférents, toxiques. (exemple de solution nutritive p. 22).
3 Méthode biochimique.
Cette méthode permet de justifier la présence d’un élément dans une solution nutritive. Elle permet aussi de voir si ces éléments sont des éléments de constitution (de composition) ou seulement des catalyseurs.
B Résultats.
On trouve une quarantaine d’éléments différents qui sont classés en trois groupes :
• Les éléments indispensables : C (45%), O (40%), H (6%), N (1%). Ces quatre éléments représentent 94% de la masse sèche de la plante. On peut y ajouter le fer (Fe) qui est indispensable dans les cytochromes.
• Les éléments macronutritifs : K (2-4%), Ca (1-2%), Mg (0,1-0,7%), S (0,1-0,6%), P (0,1-0,5%). On peut aussi trouver parmi ces éléments le sodium, le chlore et le silicium (Na, Cl, Si).
• Les oligo-éléments : Mn, Zn, Cu, B, Mo, Co, I. Les concentrations de ces éléments sont exprimées en pour-mille. Ils sont présents comme cofacteurs, de l’activité enzymatique. On trouve : K chez les algues, les épinards, les champignons et les pommes de terre ; S chez les crucifères et l’ail ; Na chez les algues marines et chez les plantes halophytes ; Si chez les graminées ; Ca est toxique chez les plantes calcifuges et est supporté chez les calcicoles ; I chez les algues marines.
Partie 1 :
NUTRITION ET METABOLISME.
Chapitre 2 : LA NUTRITION CARBONEE.
Introduction :
Dans la nature, on trouve le carbone sous deux formes assimilables :
- Le carbone minéral (CO2 ou H2CO3).
- Le carbone organique qui se trouve des les molécules organiques. On trouve deux types de végétaux :
- Ceux qui convertissent le carbone minéral en carbone organique. Ce sont les autotrophes. Ils utilisent l’énergie solaire pendant la photosynthèse grâce à la chlorophylle.
- Ceux qui n’assimilent que le carbone organique. Ce sont les hétérotrophes (c’est le cas des champignons et de quelques plantes parasites).
Remarques :
• Tous les végétaux chlorophylliens peuvent utiliser le carbone organique (souvent celui de l’urée).
• Tous les végétaux sont capables de réactions de beta-carboxylation (fixation du CO2 sur des molécules organiques). C’est une voie à faible rendement.
• Tous les autotrophes passent par un stade hétérotrophe pendant leur germination. C’est une période où l’oxygène est absent.
I Photosynthèse ou assimilation chlorophyllienne.
A Définition.
La photosynthèse est un processus physiologique par lequel les végétaux qui contiennent certains pigments (en particulier de la chlorophylle) sont capables de capter l’énergie lumineuse et de la transformer et énergie chimique (ATP et pouvoir réducteur NADPH,H+) afin de réaliser la nutrition carbonée à partir du CO2 atmosphérique, de (bi)carbonate, … Ce processus est accompagné d’un dégagement de dioxygène. Ce phénomène se déroule chez les végétaux évolués et chez les algues bleues.
B Historique.
A l’origine de la découverte de la photosynthèse, il y a les études sur les échanges gazeux.
• En 1754, Bonnet montre que les plantes dégagent de l’air «vital ».
• En 1771, Priestley découvre l’oxygène.
• En 1779, Ingen-Houz met en relation la lumière, l’oxygène et les parties vertes de la plante.
• En 1864, Sacchs montre que les parties vertes synthétisent l’amidon.
• En 1937, Hill fait la première expérience avec la chlorophylle. Il montre qu’il y a deux phases, une photochimique et une assimilatrice.
• En 1939, Willstater et Stolle établissent la formule de la chlorophylle.
• En 1954, Arnon montre que les chlorophylles réduisent le NADP et produisent de l’ATP.
• En 1955, Calvin montre que l’APG (acide phospho-glycérique) est le premier produit formé (c’est un C3).
• En 1970, Hatch met en évidence qu’une molécule en C4 pouvait être assimilée (dans le maïs).
• En 1976, Tolber met en évidence le fonctionnement des centres réactionnels (prix Nobel).
• En 1988, Mitchell et son équipe mettent en évidence les centres réactionnels des photosystèmes. Les études sur les photosystèmes peuvent se faire de différentes manières et à différents niveaux :
- Mesure des échanges gazeux
- Gain de biomasse
- Mesure de la synthèse d’amidon ou de saccharose
- Etudes de chloroplastes isolés (conversion de l’énergie lumineuse en énergie chimique).
Le chloroplaste ne suffit pas à lui tout seul pour réaliser toute la photosynthèse. La phase d’assimilation se déroule dans le chloroplaste alors que la phase d’assimilation se déroule dans d’autres compartiments cellulaires et même parfois, dans d’autres cellules.
C Le chloroplaste.
1 Structure et ultrastructure.
C’est un organite à double membrane, de forme ovoïde, de quelques dizaines de micromètres de long. Selon les espèces, on a de 10 à 100 chloroplastes par cellule (plus ils sont nombreux, plus ils sont petits). L’ensemble des chloroplastes s’appelle le plastidome qui est sensiblement constant en masse. Les chloroplastes évoluent en fonction de l’âge de la plante et de son environnement.
Les mitochondries et le péroxysome participent à la phase d’assimilation. Les chloroplastes sont généralement situés au niveau des feuilles, dans le mésophylle (ensemble de parenchymes palissadiques et lacuneux). On ne trouve jamais de chloroplastes dans l’épiderme sauf dans les stomates. Ils sont au niveau des pétioles, des tiges herbacées et de certains organes floraux. Chez chlamydomonas (algue unicellulaire), il y a un chloroplaste en forme de croissant. Chez ulothrix, il y a un chloroplaste en forme d’anneau. Chez le zygnéma, il y a deux chloroplastes en forme d’étoile. Chez spirogyne, les chloroplastes sont en forme de lames spiralées. Au cours du développement de la plante, des protoplastes se différencient en chloroplastes par des voies différentes selon les conditions externes :
- Les chloroplastes matures avec de l’amidon transitoire et un granum,
- Les chromatoplastes colorés (comme dans les fruits et les fleurs),
- Les leucoplastes où sont présentes des protéines,
- Les amiloplastes qui sont des réserves d’amidon (situés dans les feuilles et dans les racines).
Quand la cellule se divise, ses plastes vont se répartir au hasard mais ils peuvent se multiplier par divisions dans les cellules. Ces plastes sont des organites autonomes qui sont responsables de l’hérédité maternelle cytoplasmique.
Le chloroplaste possède une double membrane comme la mitochondrie. Entre ces deux membranes on trouve une phase aqueuse où il n’y a pas de réactions enzymatiques. La membrane externe est relativement perméable (elle est composée de protéines et de phospholipides). La membrane interne est imperméable : c’est une barrière sélective. Dans cette membrane, on trouve un fort taux de protéines, l’organisation moléculaire est complexe. Les lipides présents dans cette barrière sont particuliers (ce sont des galactolipides et des sulfolipides), les enzymes que l’on y trouve sont souvent des ATP-ases.
A l’intérieur du chloroplaste, on trouve le stroma (le cytoplasme) dans lequel se situe un système lamellaire formé par le développement de la membrane interne repliée sur elle-même pour former des sacs (les thylacoïdes) où sont les pigments.
Dans les thylacoïdes, l’espace est appelé lumen : c’est une phase aqueuse.
L’empilement des sacs (granum) donne une surface importante de membranes accolées qui contiennent les pigments et qui permettent une meilleure récupération de l’énergie lumineuse.
Remarque : les plantes se trouvant à l’ombre, développent fortement la surface de contact de leur granum pour avoir un apport en énergie lumineuse à peu près normal. Chez les plantes soumises à une grande luminosité, cette surface diminue.
2 Obtention de chloroplastes et réaction de Hill.
Pour isoler les chloroplastes, on réalise un fractionnement cellulaire. On part de protoplastes ou de tissus chlorophylliens. On effectue un broyage mécanique dans un milieu tamponné, avec un agent osmotique (saccharose ou manitol), des composés protecteurs (antioxydant). On travail à faible température (dans la glace) et sous lumière verte. On filtre sur une gaze, puis on centrifuge pour éliminer les débris. On a plusieurs méthodes de centrifugation :
• Centrifugation fractionnée : on réalise des centrifugations successives de plus en plus rapides (à 600 tours/mn, on enlève les parois et les noyaux ; à 1800 tours/mn, on enlève les chloroplastes et les mitochondries.
• Centrifugation sur gradient (discontinue ou non). On utilise un gradient de saccharose. On dépose l’homogénat sur le dessus puis on réalise une centrifugation de plusieurs heures. A la fin, on récupère la phase intéressante.
Une fois isolés, les chloroplastes sont capables de transformer l’énergie lumineuse en énergie chimique et de réaliser le début de la phase d’assimilation du CO2.
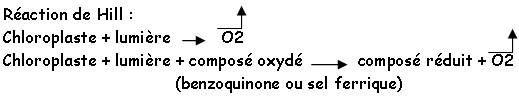
Conclusion : L’eau est la source du pouvoir réducteur et est à l’origine du dégagement d’O2.
Il y a deux phases. Une pendant laquelle l’eau est photodissociée (photochimique) et une autre pendant laquelle le CO2 est incorporé (assimilatrice).
C16O2, H218O 18O2 ; C18O2, H216O 16O2.
Réactif de Hill: tout corps pouvant être réduit par un chloroplaste isolé, en présence d’eau (in vivo, c’est le NADP).
Equation de VanNiel:
• CO2 +2*H2O [HCOH] + H2O + O2 (chez les végétaux supérieurs)
• CO2 + 2*H2S [HCOH] + H2O + S2 (chez les bactéries).
Grâce à l’action des thylacoïdes, la phase photochimique produit de l’ATP, du NADP et de un dégagement d’oxygène. Cette phase est sensible à la lumière mais insensible à la température.
La phase assimilatrice se déroule dans le stroma et exclusivement à la lumière. L’enzyme qui intervient principalement est la Rubisco qui n’est active qu’à la lumière. Une fois le CO2 fixé, le NADP et l’ATP sont utilisés.
3 Les thylacoïdes: structure et fonction.
a Composition chimique.
• Les lipides: ils représentent 30 % de la masse des thylacoïdes. On trouve 75 % de galactolipides, 15 % de phospholipides et 10 % de sulfolipides.
-Les phospholipides proviennent de glycérol estérifié en 1 et 2 par des acides gras et en 3 par un composé phosphaté.
-Les galactolipides sont estérifiés de la même façon en 1 et 2 et en 3 par du galactose.
-Les sulfolipides diffèrent par le carbone 3 du glycérol qui est estérifié par un ose sulfaté.• Les pigments: seule une partie du spectre sera utilisée 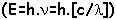 . On a de la chlorophylle (de deux types a et b) et des caroténoïdes qui sont toujours présents. On trouve des phycobillines qui sont des pigments additionnels chez certaines algues ou cyanobactéries.
. On a de la chlorophylle (de deux types a et b) et des caroténoïdes qui sont toujours présents. On trouve des phycobillines qui sont des pigments additionnels chez certaines algues ou cyanobactéries.
-La chlorophylle est un pigment vert facilement extractable par un solvant organique (elle représente 3% de la masse des chloroplastes.
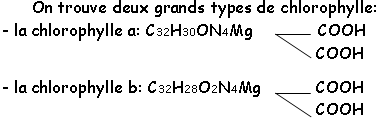
On trouve des fonctions méthyl (CH3) en 1, 3, 5, 8. En 4, on trouve une fonction éthyl (C2H5). En 2, on trouve une double liaison (CH=CH2). En 6 et 7, on trouve une fonction C2H4COOH.
Les fonctions carboxyles sont oxydées par le méthanol ou par le phytol.
La chlorophylle A à une couleur bleu-vert alors que la B à une couleur vert-jaune. Ce pigment est composé d’une queue hydrophobe et d’une tête hydrophile. Une molécule de chlorophylle est composée de 20 acides aminés. Les deux versions de la chlorophylle absorbent la lumière à différentes longueurs d’ondes : la forme a absorbe à 680 nm, alors que la forme b absorbe à 700 nm. L’absorption est réalisée grâce à l’existence de doubles liaisons conjuguées. -Les caroténoïdes: (p.4) ils ont une couleur jaune-orangé et sont des polymères de l’isoprène (C5H8). 2-méthylbut1,3diène : C=C-CH3-C=C-C ; cette forme peut-être ou non cyclisée.
Parmi les caroténoïdes, on trouve les carotènes qui sont orangés, de formule brute C40H56. Ce sont des hydrocarbures polyéniques. Ils sont présents sous trois formes  ,
,  et
et  . La forme la plus répandue est le -carotène : c’est une molécule symétrique. Le demi -carotène va former la vitamine A. Le lucopène non-cyclisé va donner le pigment rouge de la tomate.
. La forme la plus répandue est le -carotène : c’est une molécule symétrique. Le demi -carotène va former la vitamine A. Le lucopène non-cyclisé va donner le pigment rouge de la tomate.
On trouve aussi les xanthophylles qui sont des carotènes oxydés (C40H56On). Ils sont plutôt de couleur jaune. On trouve la luthéine (C40H56O2) chez les végétaux supérieurs, la fucoxanthine (C40H56O6) chez les algues, la zéaxanthine qui est un pigment du maïs.
-Les phycobillines : ces pigments sont de couleur bleue ou rouge (chez les algues). Ce sont des pigments accessoires, hydrosolubles, se comportant comme les noyaux de chlorophylle (ils ont quatre noyaux de pyrol alignés). On retrouve deux types majeurs de phycobillines : les phycoérythrobillines que l’on retrouve chez les algues rouges et qui absorbent les radiations vert-jaune, et les phycocyanobillines, possédés par les algues bleues et qui absorbent les radiations vert-orange. Selon les différents constituants, la molécule aura des propriétés d’absorption particulières. On peut dire que l’on a deux catégories de pigments :
- Les pigments primaires qui sont : la chlorophylle a chez les végétaux supérieurs. On retrouve deux types majeurs de phycobillines : les phycoérythrobillines que l’on retrouve chez les algues rouges et qui absorbent les radiations vert-jaune, et les phycocyanobillines, possédés par les algues bleues et qui absorbent les radiations vert-orange. Selon les différents constituants, la molécule aura des propriétés d’absorption particulières. On peut dire que l’on a deux catégories de pigments :
- Les pigments primaires qui sont : la chlorophylle a chez les végétaux supérieurs et un pigment bactériochimique chez les bactéries.
- Les pigments accessoires. Ce sont les autres chlorophylles, xanthophylles, phycobillines et caroténoïdes.
Le pigment primaire est le seul qui participe à la phase photochimique. Les autres pigments ne sont pas des composants de cette chaîne mais ils permettent d’augmenter l’absorption des quanta de lumière.
Ces pigments sont organisés en antennes collectrices au niveau de la membrane des thylacoïdes et leur rôle est de transférer de l’énergie vers les pigments primaires en comblant les lacunes d’absorption de ce dernier.• Les protéines : on a deux types de protéines. Des protéines réceptrices de l’énergie lumineuse qui forment des antennes et qui sont associées aux pigments. Des protéines qui servent aux transferts des électrons en formant les cytochromes. -Les cytochromes sont des chromoprotéines constituées par des métalloporphyrines. Elles possèdent un noyau tétrapyrolique (un hème) comme celui de la chlorophylle mais où le magnésium est remplacé par du fer. Les protéines qui servent à la photosynthèse ne font que transporter les électrons. Les cytochromes ont leurs six liaisons de coordination (du fer) utilisées (Fe3+ +e- Fe2+).
On trouve deux familles de cytochromes. Les cytochromes F de type C (dans les Feuilles) et les cytochromes de type B. Ce qui différencient ces cytochromes sont les substituants sur le noyau et les parties protéiques. -Les protéines Fe, S (non hémique). Leur centre actif est constitué par des atomes de fer et de soufre reliés à la partie protéique (ferrédoxine [Fd] et les protéines de Rieske). On trouve des protéines renfermant du cuivre : les plastocyanines. On trouve des complexes enzymatiques Fd, flavoprotéines (ce sont des protéines qui ont comme cofacteur FMN ou FAD) et qui fonctionnent avec du NAD ou du NADP. Il y a alors, formation d’un complexe oxydoréductase (on parle de Fd-NADP oxydoréductase).
On peut aussi trouver des protéines impliquées dans la photodissociation de l’eau (protéines mal connues). Et qui fonctionnent avec les ions Mn2+, Cu2+ et Cl-.
Il y a d’autres protéines qui sont impliquées dans la synthèse de l’ATP (facteur de couplage, ATP synthétase, ATP synthase). Ce sont des complexes enzymatiques relativement gros (visibles en microscopie électronique).
F0 est la partie qui forme le canal par lequel se déplacent les protons.
F1 est la partie enzymatique (dans le cas du chloroplaste, c’est celle qui est responsable de la production d’ATP).• Les quinones : les quinones ont un noyau benzénique et des substituants différents. Elles sont aussi des transporteurs d'électrons et de protons. Elles fonctionnent seules et n’ont pas de partie protéique. Parfois, elles n’ont pas de noyau benzénique mais de type naphtalénique.
b Les photosystèmes.
Les pigments des antennes collectrices sont sur les membranes des thylacoïdes (là où ils sont fonctionnels). L’ensemble antenne collectrice et pigment primaire (dans le centre réactionnel) constitue un photosystème (ou piège à électrons).
L’antenne sert de capteur et les pigments primaires ont le rôle de récepteurs. Les pigments des antennes absorbent les photons et les transmettent aux centres réactionnels où le pigment primaire (chlorophylle A) transforme cette énergie lumineuse en énergie chimique.
Deux photosystèmes distincts sont reliés au pigment primaire : PS I et PS II. Au niveau de l’antenne, le transfert de photons se fait de molécules à molécules. La lumière produit l’énergie d’excitation, qui de pigment en pigment, arrive au piment primaire (piège). La particularité du pigment primaire est qu’il peut expulser un ou plusieurs électrons pour les transférer à un accepteur.
Ce pigment primaire fait parti d’un système rédox, de plus, il doit retrouver son ou ses électrons pour revenir à l’état initial. Les liaisons  qui deviennent des * pour le passage à un état d’énergie supérieur. Il y plus d’énergie avec le bleu qu’avec le rouge. Pour le bleu, le passage de l’état initial au deuxième état singulé se fait par donation de chaleur. Enfin, le passage du premier état excité à l’état initial se fait par transfert d’énergie (au niveau des antennes) ou par un travail chimique (au niveau du centre réactionnel), ou bien, par une émission de lumière (fluorescence).
qui deviennent des * pour le passage à un état d’énergie supérieur. Il y plus d’énergie avec le bleu qu’avec le rouge. Pour le bleu, le passage de l’état initial au deuxième état singulé se fait par donation de chaleur. Enfin, le passage du premier état excité à l’état initial se fait par transfert d’énergie (au niveau des antennes) ou par un travail chimique (au niveau du centre réactionnel), ou bien, par une émission de lumière (fluorescence).
G = -n.FE (ne pas oublier que G<0). Il faut que le système qui va effectuer la réduction ait un potentiel plus électronégatif que le potentiel de celui qui va être réduit. Certaines réactions sont impossibles sans l’aide de l’énergie lumineuse.
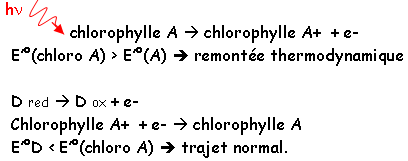
Grâce aux photosystèmes, l’énergie lumineuse permet le déplacement d’électrons.
Définition d’un photosystème : un photosystème est une unité membranaire thylacoïdale chargée de capter et de transmettre leur énergie jusqu’au centre réactionnel ou à une molécule de chlorophylle A (spéciale) afin d’expulser un électron.
Les photosystèmes ont été mis en évidence par Emerson. On a le PS I avec une chlorophylle P700 et PS II avec une chlorophylle P680 qui ont des spectres d’absorption différents.
Le photosystème II est associé à la photodissociation de l’eau alors que le PS I est associé à la réduction de Fd et du NADP.
Ces deux photosystèmes sont capables de prélever du pouvoir réducteur dans la molécule d’eau. (E’°= 810mV).
c Trajets des électrons au cours des « réactions claires ».
On a deux types de trajets : cycliques et non cycliques.
• Le trajet non cyclique :
Eau PS II PS I NADP
On aura, soit le trajet normal, soit le trajet thermodynamique.
Photodissociation de l’eau :
- H2O 4 H+ + 4HO-
- 4 HO- 4 e- + 2 H2O + O2
2 H2O 4H+ + O2 + 4 e-
Bilan :
• un O2 émis pour 2 NADP réduits
• remontée thermodynamique de 2*4 e- = 8 photons
• exigence quantique = 8
• rendement quantique = nombre d’O2 dégagé(s) par les quanta absorbés = 1/8
Par la phosphorylation, on obtient +2 ATP (c’est la somme des ATP couplés au transfert des électrons activés par la lumière).
• Le trajet cyclique :
Plastocyanine PS I 20 chloro-13-ydroxychlorophylle PS I
remontée thermodynamique de deux électrons
+1 ATP par la phosphorylation.
Ce trajet se réalise en parallèle au trajet non cyclique et apporte un complément d’ATP.
Trajet non cyclique : A= phéophytine
D= H2O
Trajet cyclique : D= plastocyanine
A= 20 chloro-13-hydroxychlorophylle
• Couplage chimio-osmotique :
-La synthèse d’ATP et le transfert d’électrons sont couplés chimiosmotiquement. Pendant la photosynthèse, les transferts des électrons au niveau de la membrane sont à l’origine d’un gradient électrochimique de protons (µ). Ce gradient est compensé par les facteurs de couplage des thylacoïdes, ce qui entraîne la production d’ATP.
-On obtient une différence de concentration de protons de part et d’autre de la membrane thylacoïdale. Au niveau stromatique, le milieu devient plus alcalin (pH=8), alors que du côté du Lumen, le pH est de 5.
Les complexes d’ATP synthase peuvent fonctionner dans les deux sens.
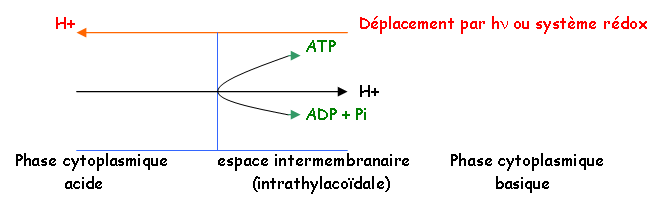
Le retour des protons permet la récupération d’énergie. La phosphorylation réalisée dans le chloroplaste est 10 fois supérieure à l’énergie produite par la mitochondrie.
Trajet non cyclique :
8 photons 2 NADPH,H+ + 2 ATP + 2 x ½O2
Trajet cyclique :
2 photons 1 ATP
Les produits de ces deux réactions vont permettre l’intégration d’un CO2 Par exemple, dans un champs de tournesol, la production de dioxygène est de 180 kg/ha/h.
4 Le stroma : la Rubisco et le cycle de Calvin.
L’ATP est libéré dans le stroma. On trouve dans celui-ci, toutes les enzymes nécessaires au métabolisme ainsi que tous les métabolites. Une partie de la phase d’assimilation du CO2 se déroule dans le stroma. Le premier produit formé est l’Acide PhosphoGlycérique (APG).
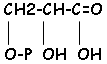
L’élévation de la quantité d’APG correspond à la diminution du RubP (ribulose 1,5 diphosphate). CH2O(PO32-)-C(=O)-CH2-CH2-CH2O(PO32-) Il y a ensuite, mise en évidence de l’enzyme responsable : la RubP carboxylase. (C5+CO2C62 C3 ou 2 APG). C’est la première phase de la fixation de la photosynthèse qui est rendue possible grâce à la fonction carboxylasique de la RubP (la Rubisco a deux activités : carboxylase et oxygénase).
Dans les réactions qui vont suivre, le carbone va être assimilé puis incorporé dans des molécules d’oses phosphates. Cet ensemble de réactions constitue le cycle de Calvin (cycle réducteur se déroulant en trois phases).
Phase 1 : carboxylation de RubP APG (fixation de CO2)
Phase 2 : réduction des APG trioses-P (réduction du carbone fixé)
Phase 3 : régénération du RubP
La phase 2 est une réduction de l’APG en triose. La phosphorylation est réalisée par une kinase (à partir d’ATP). Quand cette réaction a lieu à partir de Pi, l’enzyme est une phosphorylase. On obtient la forme 1,3diphoshoglycérique, puis, la déshydrogénase (qui fonctionne avec du NADP) va former le glyceraldéhyde 1,3diphosphate qui est en équilibre avec la forme cétose (CH2OP-CO-CH2OP). Le passage de l’une à l’autre des deux formes est réalisée par une isomérase. Ces deux formes forment les trioses phosphates.
Pendant la phase 3 (de régénération), la Rubisco est réglée par la lumière (elle ne fonctionne que le jour). Un des six trioses phosphates part vers la synthèse de matière organique.
5 trioses phosphates donnent 3 pentoses phosphates
I. C3 + C3 C6 (aldolase)
II. C3 + C6 C5 +C4 (trancétolase)
III. C3 + C4 C7 (aldolase)
IV. C3 + C7 C5 + C5
Le ribose est transformé en ribulose par une isomérase.
Les hexoses primaires sont les produits terminaux de l’assimilation du CO2 pendant la photosynthèse. Un fructose phosphate se forme à partir de 6 CO2 et de 6 RubP.

La Rubisco est un ensemble de deux sous-unités (A et B), répétée chacune, 8 fois. Elle résulte de l’activité des deux génomes (cellulaire et chloroplastique).
La grosse sous-unité A (55 kDa) est formée dans le chloroplaste et sert de site catalytique.
La petite sous-unité B (15 kDa) est sous la forme de précurseurs synthétisés dans le cytoplasme puis qui pénètrent dans le chloroplaste.
A8B8 a un poids moléculaire de 560 kDa et représente jusqu’à 50 % des protéines solubles d’une feuille. [CO2]air (0,03%) correspond à 10 µmol. Depuis 1971, on sait que cette enzyme permet la fixation d’O2.
C5 + O2 [C5] C2 + C3 APG (C2 est l’Acide PhosphoGlycolique). C’est le début de la photorespiration : c’est un processus biochimique qui s’associe au cycle de Calvin (dans le stroma).
Remarque : pour que la photosynthèse fonctionne, il faut que l’affinité de la Rubisco pour le CO2 soit beaucoup plus grande que l’affinité pour O2. (Si l’affinité augmente, le Km diminue).
D Les plantes de type photosynthétique C3.
Les deux activités de la Rubisco s’expriment en même temps et s’expriment en fonction des quantités d’O2 et de CO2. On a, à la fois, les cycles de Calvin et la photorespiration.
• Activité de la carboxylase
2 C3 donnent 2 acides phosphoglycériques par le cycle de Calvin.
• Activité de l’oxygénase
1 C3 par le cycle de Calvin
1 C2 par la photorespiration
Au cours du cycle photorespiratoire, deux acides phosphoglycoliques (des C2) vont sortir du chloroplaste, subir le cycle photorespiratoire et à la fin de celui-ci, on obtient un C3 (APG) qui va être récupéré par le chloroplaste (c’est le cycle de Tolber).
On tient compte des mouvements gazeux pour pouvoir quantifier la photosynthèse. Le taux d’assimilation est équivalent à la photosynthèse nette (PN) et la capacité d’assimilation est identique à la photosynthèse brute (PB).
PN = PB – R. On peut donc dire que les végétaux poussent plus vite quand leur respiration est faible. Ce cycle en C3 existe chez la majorité des plantes.
Bilan de la photorespiration :
Il y a consommation d’O2 (Rubisco et glycolate oxydase) et dégagement de CO2 (passage de 2 acides phosphoglycoliques à un APG).
- On a l’élimination de l’acide phosphoglycolique, qui est toxique pour la plante, par la photorespiration.
- Elle permet la biosynthèse d’acides aminés (protéogenèse).
- Biosynthèse d’acide glycérique qui va alimenter le cycle de Calvin.
- Biosynthèse de NADPH,H+ dans la mitochondrie (phosphorylation oxydative).
- Elle permet le fonctionnement du cycle de Calvin avec les stomates fermés.
- On assiste à la perte de CO2, d’où une diminution de la photosynthèse nette chez les plantes en C3.
- Il y a formation de NH4+ à partir des fonctions amines, qui, pour sa réassimilation demande de l’ATP et du pouvoir réducteur.
E Les plantes de types photosynthétiques C4.
Chez le maïs, on trouve un produit stable en C4 : le malate (COOH-CHOH-CH2-COOH). C’est un produit qui dérive de l’oxaloacétate (COOH-C=O-CH2-COOH). On a le même résultat chez les graminées, les monocotylédones et chez quelques dicotylédones comme l’amarnthe et l’euphorbe.
Ces plantes on une anatomie foliaire particulière : c’est une structure en anneau ou en couronne. Le parenchyme médulaire (mésophylle) est en contact avec les gaz. Autour des vaisseaux, on trouve des cellules spécialisées qui forment la gaine périvasculaire. Ces cellules sont fortement accolées.
Ces tissus particuliers ont des fonctions particulières : les cellules du mésophylle sont en contact avec le CO2 atmosphérique et c’est à leur niveau qu’a lieu la fixation primaire du CO2. On assiste à une double carboxylation séparée dans l’espace.
|
Mésophylle
Chloroplastes granaires
PS I et PS II
CO2 atmosphérique
PEP Carboxylase
|
Cellules de la gaine
Chloroplastes agranaires
(pas d’empilement de granum)
PS I
CO2 malique
Rubisco
|
Conséquence : les deux types de cellules vont absorber la lumière et donc, avoir une phase photochimique, mais, pour les cellules de la gaine, il n’y a pas de PS II, ce qui empêche la phase acyclique d’où l’absence de pouvoir réducteur.
Le mésophylle : il y a fixation du CO2 atmosphérique par le phosphoénolpyruvate carboxylase (PEP) qui est une enzyme cytoplasmique à très forte affinité pour le CO2. Cette affinité entraîne l’existence de cellules avec des chloroplastes sans Rubisco ; phénomène qui lui, entraîne le transport de l’acide malique vers les cellules de la gaine.
La gaine : dans le stroma, on note la présence de Rubisco, mais éloignée de l’atmosphère. Cet éloignement oblige la fixation de CO2 (malique) par décarboxylation de l’acide malique.
AOA = Acide Oxalo Acétique.
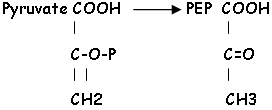
L’enzyme malate déshydrogénase est NADP dépendante. Elle se trouve dans la gaine où le malate est décarboxylé et déshydrogéné par l’enzyme malique. Malate CO2 + Pyruvate + NADPH
Le pyruvate est phosphorylé par la pyruvate phosphate dikinase dans le mésophylle. Le CO2 qui retourne dans la gaine est pris par la Rubisco pour faire tourner le cycle de Calvin, puis celui de Tolbert.
Conséquences : - les plantes en C4 fixent plus de CO2 que les C3, donc, leur production de biomasse est plus importante. Cette supériorité est expliquée par l’activité de la PEP carboxylase, par le système de transport du malate vers les cellules de la gaine qui entraîne une augmentation de concentration du CO2 dans les chloroplastes des cellules de la gaine. Ici, la concentration en CO2 des cellules de la gaine est vingt fois supérieures à la concentration de l’atmosphère en CO2.
- La diminution de l’activité oxygénante de la Rubisco entraîne une faible photorespiration. De plus, le CO2 libéré par cette respiration doit traverser les cellules du mésophylle par la PEP carboxylase. On obtient chez les C4, une photosynthèse nette égale à la photosynthèse brute.
- Les C4 utilisent la lumière forte, ce qui leur permet d’importantes synthèses en ATP et NADP.
- Le système de fixation contribue à économiser l’eau. Ces plantes peuvent donc maintenir leurs stomates fermés afin de diminuer l’évapotranspiration. L’efficacité est suffisante, car il y a assez de CO2 pénétrant qui est fixé par la PEP.
Remarque : Si les C4, en plein soleil et avec beaucoup d’eau ont une forte productivité, ils colonisent en général des milieux fortement exposés au soleil mais faibles en eau. Les C4 sont toujours des angiospermes.
F Les plantes de types photosynthétiques CAM.
CAM : Métabolisme Acide Crassulacéens.
On trouve ce type photosynthétique chez les cactacées et chez quelques liliacées et broméliacées (plantes des régions arides). Ces plantes ferment leurs stomates le jour, pour éviter les pertes d’eau. Elles sont donc obliger de fixer le CO2 la nuit alors que la Rubisco ne fonctionne pas. On assiste au développement d’un mode de fixation, comme chez les C4, sur une double carboxylation. Toutefois les deux carbones sont séparés dans le temps et non dans l’espace.
Pendant la phase nocturne, La PEP carboxylase fixe le CO2 sur l’AOA. Cette enzyme fonctionne dans le cytoplasme et avec du NAD. Il y a accumulation d’acide malique dans la vacuole (par un transport actif). Le pH de la vacuole diminue et se stabilise entre 3 et 4.
Le jour, l’acide malique sort de la vacuole par un transport passif (le pH de la vacuole remonte jusqu’à 6). Le malate repris par l’enzyme malique (dans le chloroplaste) libère des CO2 qui vont rentrer dans le cycle de Calvin grâce à la Rubisco. Il y a aussi photorespiration.
L’accumulation d’acide malique a ses limites (l’espace est limité). Le PEP vient de la dégradation d’amidon, formé pendant la photosynthèse le jour.
Il y a des problèmes de régulation pour les enzymes. Ici, tout se passe dans la même cellule. Il faut tout réguler pendant la journée. Pendant le jour, la PEP carboxylase est inhibée par le malate. C’est une adaptation physiologique à la faible disponibilité en eau. Le cycle de Calvin est indépendant du CO2 atmosphérique.
L’augmentation de la concentration en acide malique entraîne l’augmentation de la pression osmotique qui provoque une attraction pour l’eau (notamment, l’eau atmosphérique). Il y a une importante absorption de rosée.
On retrouve le système CAM chez certaines fougères. C’est une adaptation physiologique non-évolutive. Dans ce cas, on retrouve un peu de photorespiration qui reste faible, car il y a peu d’eau.
C3 : 1 CO2 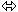 3 ATP et 2 NADPH,H+
3 ATP et 2 NADPH,H+
C4 : 1 CO2 5 ATP et 1 NADPH,H+
G Biosynthèse de l’amidon et du saccharose.
C’est l’utilisation du sixième triose du cycle de Calvin.
Chez les C3, C4 et CAM, le sixième triose part former du fructose qui donnera des polysaccharides. En général, les glucides formés sont de l’amidon (pour le stockage) ou du saccharose pour le transport.
L’amidon :
• C’est un glucane avec des liaisons en 16 et 14.
• L’amylase coupe les liaisons en D glucane 14.
• Les amilopectines : Coupent les liaisons des chaînes ramifiées en 14 et 16.
• La synthèse de l’amidon se fait par l’amidon-synthase associé à l’enzyme Q.
• La dégradation de l’amidon se fait par l’amidon phosphorylase associé à l’enzyme D. Pendant la germination, on peut avoir l’ et amylase, l’isoamylase est associée à l’enzyme R.
Glucose : CHO-(CHOH)4-CH2OH ; Fructose CHO-C=O-(CHOH)3-CH2OH.
L’amylose est facilement soluble. L’amidon, lui, n’est pas soluble ; il s’accumule toute la journée dans les chloroplastes (? amidon transitoire) et est rapidement utilisé. S’il est synthétisé en grande quantité, il sera stocké dans les amyloplastes.
Le saccharose est soluble. C’est une association de glucose et de fructose.
Pour pouvoir être utilisé, l’amidon est transformé en saccharose pour son transport (par la sève élaborée). Pour être synthétisé dans les chloroplastes, l’amidon demande l’utilisation d’ATP. Le saccharose synthétisé dans le cytoplasme demande de l’énergie qui vient de l’UTP.
P. 14, la synthèse du saccharose : il y a transport du triose à travers la membrane chloroplastique sous la forme DiHydroAcétonePhosphate (DHAP). Le passage de la membrane demande une entrée de Pi dans le chloroplaste.
4 DHAP Fructose
(4) 2 DHAP (2) 1 Fructose 1,6 BP
Il y a ensuite déphosphorylation du fructose BP qui donne du fructose 6P. Quand on a deux fructose 6P, un reste fructose et l’autre passe sous la forme de glucose 6P grâce à une isomérase.
Glucose 6P glucose 1P grâce à une phosphoglucomutase. Le glucose 1P est activé grâce à l’UDP en donnant de l’UDP-glucose (l’enzyme est une uridyltransférase).
UDG-gluc + Fr6P saccharose P grâce à une saccharose synthase.
Saccharose P Saccharose + Pi (grâce à l’action d’une saccharose 6phosphatase). Le saccharose va rejoindre les vaisseaux du phloëme.
L’amidon est formé dans le chloroplaste :
2 trioses P 1 Fructose1,6BP Glucose 1P.
La régulation de la synthèse de l’amidon et du saccharose :
Quand il y a beaucoup de trioses dans le chloroplaste, il y a activation de la formation d’amidon. Quand il y a beaucoup de Pi dans le chloroplaste, c’est le signe d’un besoin en saccharose qui inhibe la voie de synthèse de l’amidon.
H La photosynthèse au niveau de la plante.
Les manifestations les plus visibles de la photosynthèse sont une augmentation de biomasse (matière organique) et une augmentation des échanges gazeux (O2/CO2).
1 Intensité de la photosynthèse.
On mesure l’augmentation de la biomasse et les concentrations en glucides, lipides et protides. Par le calcul, on voit la quantité de CO2 nécessaire à cette augmentation de biomasse.
Le plus simple est de mesurer les échanges gazeux par différentes méthodes (appareils manométriques de Warburg, analyseur de gaz à infrarouge, dosage de produits radioactifs). La quantité de CO2 est la résultante du CO2 absorbé moins la quantité de CO2 émis : c’est la mesure de la photosynthèse nette. L’intensité de la photosynthèse est exprimée en µmol ou mmol par surface assimilatrice et par unité de temps.
Pour les C3, on a 20 mg de CO2 /dm²/h.
Pour les C4, on a 40 mg de CO2 /dm²/h.
Pour les CAM, on a 8 à 10 mg de CO2 /dm²/h.
Un champs de blé fabrique 10 tonnes de matière sèche/ha/an.
Un champs de maïs fabrique 25 tonnes de matière sèche/ha/an.
Grâce à la photosynthèse, les végétaux élaborent 100 milliards de tonnes de biomasse. Cette quantité de biomasse représente 150 milliards de tonnes de CO2, d’où l’importance des hétérotrophes et leur production de CO2. Toutefois, le rendement de la photosynthèse est relativement simple.

2 Influence des facteurs du milieu.
Sur une plante, chaque fonction physiologique va être influencée par différents facteurs. En fonction de ces facteurs, va s’appliquer la loi des facteurs limitants de Liebig : l’intensité d’une fonction qui dépend de plusieurs facteurs n’est augmentée que par le facteur qui se trouve au niveau le plus faible car c’est un facteur limitant.
La photosynthèse est limitée par la lumière, la concentration en CO2, la température, la concentration en oxygène, l’eau, la pollution et l’alimentation minérale.
a L'éclairement.
C’est le flux énergétique, transporté par la lumière et reçu par unité de surface (il s’exprime en W/m²). L’éclairement solaire maximum représente 950W/m², mais seulement 54% se trouve dans le visible (soit 510W/m²). Les radiations actives pour la photosynthèse (PAR) sont entre 400 et 700nm. On parle de flux de photons, 510 W/m² représentent 1800µeinstein/m²/s. L’œil évalue l’intensité lumineuse qui est exprimée en Lux. 510W/m² 100.000Lux.
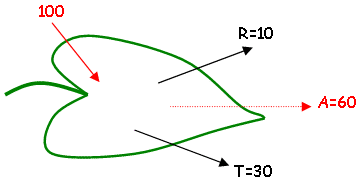
Au niveau de la feuille : A=60=40 (évaporation) + 19 (chaleur) + 1 (photosynthèse).
La quantité d’éclairement intervient et les plantes comme les sciaphilles (plantes d’ombres) vont accepter un faible éclairement. Les plantes héliophiles vont demander un éclairement plus important. Les plantes d’ombre ont des feuilles peu épaisses avec peu de parenchymes. Toutefois, les chloroplastes sont pourvus de nombreux thylacoïdes qui leur permettent de compenser ce manque de lumière.
Le point de compensation lumineux est la valeur de l’éclairement pour laquelle la photosynthèse nette est nulle (donc photosynthèse brute = photorespiration). C’est seulement au-dessus de cette valeur que la plante va croître. On obtient des courbes de saturation différentes selon le type de plantes :
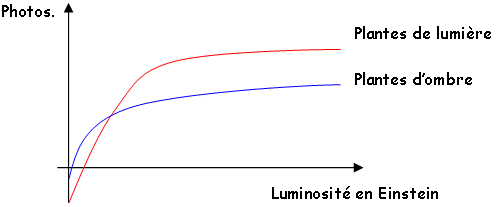
Les plantes en C3 arrivent à saturation au tiers du plein soleil. Les C4 ne sont pas gênés par une intensité lumineuse maximale. La qualité de la lumière joue un rôle dans l’assimilation. Dans certaines serres, on ajoute certaines radiations pour améliorer les cultures.
b la teneur en CO2.
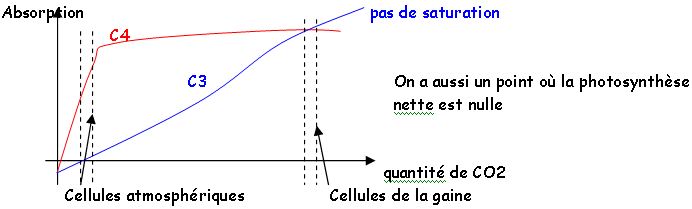
La concentration de l’atmosphère en CO2 est de 0,03%.
Artificiellement, on peut augmenter la teneur en CO2 jusqu’à 1% (au-dessus, la concentration devient toxique). C’est le principal facteur limitant de la croissance des plantes. A température élevée, les stomates se ferment, empêchant donc l’entrée du CO2. Les stomates des plantes sont ouverts le jour et fermés la nuit (inversement pour les CAM). Si on mesure l’assimilation en fonction du CO2, on a aussi des courbes de saturation.
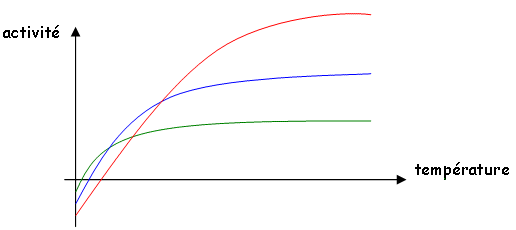
c La température.
La température agit sur les réactions enzymatiques (sur la phase assimilatrice). La réaction photochimique est sensible la lumière alors que les réactions enzymatiques sont sensibles à la température.
d la teneur en O2.
La concentration moyenne en O2 de l’atmosphère est de 21%.
Cette concentration est néfaste pour les C3 car elle va faire photorespirer les plantes. Le déficit hydrique fait diminuer la photosynthèse à cause de la fermeture des stomates (il y a toujours un fort déficit en eau).
e Les autres facteurs.
• La nutrition minérale : c’est le besoin de minéraux essentiels comme Mg, Fe, …
• Les facteurs de pollution : les oxydes vont inhiber des enzymes ou dégrader la chlorophylle.
II Hétérotrophie par rapport au carbone, Conclusion.
Dans le cas des végétaux incapables d’utiliser l’énergie lumineuse pour réduire l’oxygène, l’énergie provient de la matière organique (du catabolisme). Ces plantes sont obligées d’utiliser des composés carbonés (le plus souvent, ce sont des glucides, mais les lipides et protides peuvent aussi être utilisés). Pour pouvoir absorber cette matière organique, le végétale doit avoir à sa surface, des molécules capables de fixer les substrats.
Ces organismes doivent être capables d’excréter des enzymes qui vont pré-dégrader la matière organique. Il faut que les substrats puissent pénétrer dans l’organisme hétérotrophe. Une fois dans l’organisme, les substrats doivent être entièrement dégradés.
On a différents types d’hétérotrophes :
A Les saprophytes.
Ce sont des organismes qui vivent au dépend de la matière morte. On les trouve au niveau du sol, sur des végétaux morts (ils vont transformer la matière organique en humus). Ils participent au recyclage des éléments minéraux. Ce sont essentiellement des mycètes.
B Les parasites.
Ce sont des organismes qui vivent au dépend de végétaux vivants (les hôtes). Ils sont en général à l’origine de maladies chez cet hôte. On trouve :
• Les parasites obligatoires.
Ce sont les champignons qui ne peuvent se développer que sur un hôte vivant et qui est spécifique d’une espèce ou d’une variété. On ne sait pas bien les cultiver in vitro. On trouve par exemple Puccinia graminis qui est la rouille du blé (et de l’orge)(voir le cours de BV), Ustilgo maydes qui donne le charbon du maïs.
• Les parasites facultatifs.
Ce sont soit des saprophytes, soit des parasites qui « attaquent » les végétaux. On trouve, par exemple, Colletotrichum lindemuthianum qui provoque l’anthracnose (pourriture sèche) chez le haricot, Phytophthera qui est une famille de champignons qui attaquent le tabac et l’horticulture.
• Les angiospermes parasites obligatoires :
- Les haloparasites (plantes parasites de plantes) : ce sont des plantes qui ne synthétisent pas de chlorophylle. On trouve la cuscute qui se développe sur les parties aériennes en formant des réseaux de filaments blancs. On a aussi l’orobanche qui se fixe sur les racines en formant des fleurs brun/rouge.
- Les hémiparasites : ils sont autotrophes pour le carbone, mais utilisent l’hôte pour lui prendre l’eau et les sels minéraux. On trouve Viseum album (le gui) qui produit des graines collantes, qui, sur l’arbre, s’insèrent dans la branche et détournent la sève brute.
C Les symbiotiques.
Symbiose : association à bénéfices réciproques (vieille définition). On trouve différents types de symbioses :
• La symbiose lychénique : c’est une association de Mycobiontes (mycètes) et de photobiontes (algues) ou cyanobactéries. En général, ces espèces peuvent vivre seules, mais ils s’associent pour former les lychens. Leurs relations métaboliques sont mal connues. Les algues amèneraient les produits de la photosynthèse (il y association quand les conditions deviennent difficiles). Les lychens sont des formes très résistantes aux conditions extérieures (eau et température). Ils sont sensibles aux pollutions (surtout au SO2).
• Les symbioses mycorhiziennes : ce sont des associations de champignons et de racines de cormophytes. C’est la symbiose la plus répandue dans la nature mais la plus difficile à étudier. Les champignons assurent la fonction de poils absorbants (pour l’eau et les sels minéraux). Les Ectomycorhizes sont des champignons dont les hyphes restent à l’extérieur des cellules racinaires (le mycélium se développe entre ces cellules). On trouve les ascomycètes (Tuber melanosporum : truffe) qui mycorhizent le chêne, les basidiomycètes (comme le bolet) mycorhizent les chênes et les pins, les Endomycorhizes sont des mycètes qui envoient des suçoirs dans les cellules de l’hôte.
• Les symbioses bactériennes : certaines bactéries (parmi, les cyanobactéries, les actinomycètes, les rhizobiacées) vont s’associer à des plantes et fixer l’azote atmosphérique (on parle d’engrais « verts »). Les cyanobactéries ont peu d’organisation, elles forment des amas cellulaires. Les actinomycètes forment des nodules (excroissances souvent au niveau aérien) où la bactérie transforme l’azote. On trouve dans cette dernière classe, Frankia qui s'associe aux arbres des pays chauds comme le Sahel. Rhizobium affecte uniquement les légumineuses.
Conclusion :
On pourrait penser que les hétérotrophes sont des parasites pour les autotrophes, mais en fait, ce sont des formes complémentaires.
Les autotrophes sont appelés « producteurs primaires », ils sont mangés par les herbivores, eux-mêmes, mangés par les carnivores puis par d’autres carnivores… Les déchets sont recyclés par les bactéries et les mycètes. Ces éléments retraités sont repris par les producteurs primaires (recyclage de l’O2 et du CO2).
Partie 1 :
NUTRITION ET METABOLISME.
Chapitre 3 : LA NUTRITION AZOTEE.
A Les différentes formes de l’azote présent dans le minéral.
L’azote sous forme organique ou minérale représente 1 à 5% de la matière sèche. On trouve l’azote dans les protéines qui sont caractérisées par un taux moyen de 16% de cet élément. Le dosage de l’azote permet un dosage des protéines. Le pourcentage de protéines est : %N*6,25 (6,25 représente le coefficient de Kjeldahl : 100/16=6,25). C’est la méthode la plus précise mais pas la plus utilisée. On trouve l’azote dans les acides nucléiques, les coenzymes, les vitamines, les hormones… Quand l’azote est sous forme minérale, c’est sous une forme ionique comme NH4+ ou NO3-.
B Les différentes formes d’azote disponible dans la biosphère.
1 L’azote atmosphérique.
Il représente 78% de l’air, c’est donc la principale source. Toutefois, seules quelques plantes qui vivent en symbiose (bactéries ou algues) sont capables d’utiliser directement l’azote atmosphérique.
2 L’azote du sol.
L’azote a cinq électrons sur sa couche externe dont trois sont célibataires et peuvent donc établir des liaisons covalentes. Le nombre d’oxydation de l’azote varie de –3 à +5.
- On trouve l’azote minéral sous trois formes : NO3-, NO2-, NH4+.
- L’azote organique se trouve dans des molécules complexes qui peuvent être des protéines ou des acides aminés. Ces molécules se trouvent surtout au niveau de l’humus. La matière organique décomposée va être à l’origine de l’azote utilisé par la plante.
a Transformation rapide, décomposition, humidification, minéralisation.
On trouve différents types de décomposition :
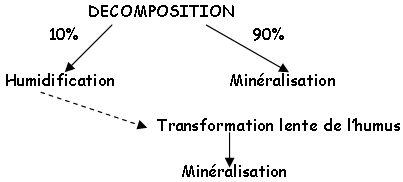
DECOMPOSITION
10% 90%
La transformation rapide nécessite un processus microbien puis une phase physico-chimique (cette phase entraînera la libération de substances nutritives). Certains composés (10%) sont difficilement dégradables et passent donc par une phase de dégradation lente.
b La transformation lente.
Elle a surtout lieu pour les composés comme la cellulose, la lignine. Ces composés sont lentement oxydés, polymérisés (processus chimique). Il y a alternance de dessiccation et d’humidification et intervention de micro-organismes (grosses molécules très grosses molécules minéraux). L’humus est une fraction colloïdale de la matière organique obtenue par synthèse microbienne (physico-chimique) à partir de matière organique végétale.
c La minéralisation.
• La protéolyse : les protéines sont dégradées en acides aminés ou en amides.
• Ammonisation : l’urée va donner de l'ammoniac, des sels ammoniacaux, des carbonates d’ammonium (CO3(NH4)2). Les sels d’ammonium vont subir la nitrification.
• La nitrification transforme l’ammoniac et le NH4+ en ions nitrate par 2 oxydations successives. NH4+ (-III) NO3- (+V). La somme des nombres d'oxydation donne la charge de l’ion.
• La nitrosation : l’acide nitreux et le nitrite. 2 NH4+ + 3 O2 2 NO2- +2 H2O + 4H+ (G’°=-542kJ/mol). Il y a production d'énergie. La réaction et réalisé par les bactéries au niveau du sol. Ces bactéries sont du genre Nitrosomonas. Les ions NO2- sont des ions toxiques.
• La nitratation : l’acide nitrique, le nitrate. 2 NO2- + O2 2 NO3- (G’°=-155kJ/mol). Cette réaction est réalisée par les bactéries du genre Nitrobacter qui sont des chimiolithotrophes. Ces bactéries sont autotrophes pour le carbone et utilisent l'énergie pour assimiler ce carbone.
C Assimilation de l’azote minéral par la plante.
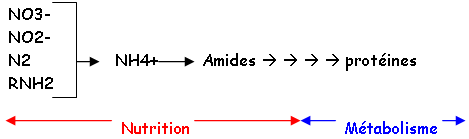
Quel que soit l’ion de départ, il passe toujours par le stade NH4+. Bien que les végétaux puissent utiliser l'ammoniaque, ils se servent surtout de l’ion NO3- : c'est la réduction des nitrates. L'assimilation de l'azote minéral est réalisée dans les racines, dans les bourgeons ou dans les feuilles.
Dans les racines, NO3- donne des amides et des uréides transformés puis transportés par le xylème.
Dans les tiges et les feuilles, NO3- donne des amides qui se sont transportés par le phloëme. Les enzymes sont : la nitrate réductase (NO3- NO2- 2e-) ou la nitrite réductase (NO2- NH4+ 6e-).
1 Assimilation des nitrates.
a La nitrate réductase. NO3- ? NO2- (2e-).
Cette enzyme est une flavo-métallo-protéine cytoplasmique. E’°= +0,46. Dans la racine, il y a utilisation de NADPH ou de NADH. Cette réaction inhibée par le NH4+ et a des effets inducteurs par le NO3-. L’enzyme est sensible à la lumière et aux hormones (cytokinines).
b La nitrite réductase. NO2- ? NH4+ (6e-).
L’enzyme est une métallo-protéine qui renferme du fer dont le rôle est de catalyser une série de réactions dont l'intermédiaire est mal connu. Dans le stroma, il y oxydoréduction avec le donneur (ferrédoxine ou NADPH). Les plantes préfèrent absorber l’ion NO3-.
2Assimilation de l'azote.
On a deux possibilités majeures :
a La glutamate déshydrogénase (GDH).
Il existe du GDH mitochondrial et chloroplastique. Ce produit se transforme :
cétoglutamate + NH4+ Glutamate
COOH-C=O-CH2-CH2-COOH //// COOH-CNH2-CH2-CH2-COOH
C'est le phénomène d’amination réductrice. L’enzyme fonctionne avec le NADPH ou le NADH selon sa localisation.
b La glutamine synthase (GS-GOGAT).
GS : glutamine synthétase ; GOGAT : glutamate synthase.
glutamate + NH4+ glutamine
glutamine + cétoglutarate glutamate + glutamate.
Le Glutamate va vers le métabolisme.
COOH-CNH2-CH2-CH2-C(=O)-NH2 : formule de la glutamine.
La première en réaction demande l'utilisation d’ATP. Il est sous sa forme cyclique, dans le chloroplaste et le cytoplasme de tous les végétaux supérieurs. Dans les nodules (dans le cas des symbiontes) et chez les cyanobactéries, on a, comme pour le GDH, des iso-enzymes.
La réaction de trans-amination (à partir de la glutamine et du glutamate) donnera toujours des amides, qui, celle-ci, cède un ammonium à des acides cétoniques. On a alors différentes familles d’acides aminés :
- glutamate : 5C
- aspartate : 4C (il vient de l’AOA). AOA ? CCO-CNH2-CH2-COOH
- Pyruvate : 3C. Pyruvate Alanine (COOH-C(NH2)-CH3).
- Sérine : CHOH à la place de l’alanine.
- Shikimate : 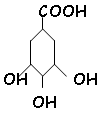
Cette forme est à l’origine de tous les AA aromatiques.
D Utilisation de l’azote atmosphérique.
Les plantes utilisant cet azote sont dites diazotrophes. Elles possèdent une enzyme particulière (la nitrogénase). C'est une enzyme de micro-organismes.
1 Les différents types de micro-organismes fixateurs d'azote.
- les fixateurs libres : Ils vivent dans le sol et quand ils meurent, ils donnent l’humus. Cet humus sera utilisé par la plante.
• Les hétérotrophes au carbone : On trouve les bactéries aérobies comme Azobacter. On trouve aussi des bactéries anaérobies de type Clostridium.
• Les autotrophes au carbone : on trouve des photo-bactéries anaérobies a un photosystème comme celles de type Rhodospirillum. On peut aussi trouver des cyanobactéries comme nostoc et Anaboena.
Azospiryllum est un fixateur libre de l'azote qui vit dans la rhizosphère des pieds de maïs.
- Les fixateurs symbiotiques. On trouve les hétérotrophes au carbone comme, les rhizobiacées (anaérobies), par exemple, les légumineuses. On pouvait aussi, trouver des actinomycètes (frankia, qui parasite les plantes ligneuses). Parmi les autres opprobres au carbone, on trouve au des cyanobactéries comme nostoc Anaboena mais aussi des hépatiques, des fougères, cycadacées, gunnera...
Définition d’un nodule : c'est un organe différencié dans la plante, où se développent les bactéries fixatrices d'azote. Les bactéries se transforment en bactéroïdes (qui fixent l'azote) de façon irréversible.
2/ La nitrogénase (ATP dépendante).
Cette enzyme est composée de six sous-unités. Ces six parties forment une protéine ferrique à deux sous-unités (dinitrogénase réductase azoferredoxine). C’est elle qui utilise l'ATP et qui transfert les électrons à l'autre sous-unité : la dinitrogénase (protéine qui contient du fer-molybdène). Elle qui fixe l'azote en formant l'ion NH4+. Elle reçoit les électrons de la première partie protéique et fixe l'azote. Le résultat de cette activité représente 10% des protéines solubles dans les bactéroïdes.
Page numéro 16 : La réduction de l’azote atmosphérique.
Le pouvoir réducteur vient de la ferrédoxine ou d'autres réactions comme la décarboxylation oxydative du pyruvate, cette réaction demande de l'ATP et produit de l'hydrogène. L'ATP vient de la photosynthèse ou de la phosphorylation oxydative. L’azote est transformé en matières organiques comment précédemment.
Le problème de la régulation : la dinitrogénase est très sensible à l'oxygène et est inhibée irréversiblement par des traces d'oxygène. Dans le cas des organismes anaérobies, il n'y a pas de problème.
Chez les algues bleues, la compartimentation de la nitrogénase, dans les hétérocystes empêche ce problème (photosystèmes).
Dans le cas des fixateurs symbiotiques, libres et aérobies, l'oxygène est piégé par une respiration amplifiée. Dans le cas particulier des symbiotiques, on a en plus des métallo-protéines (chromoprotéines) qui contiennent un hème : la leghémoglobine qui piège l'oxygène. Cette protéine représente 40% des protéines des nodules de légumineuses. La leghémoglobine fait partie des nodulines, cette protéine est synthétisée par la plante seulement dans les nodules. L'azote ammoniacal et les nitrates sont des répresseurs pour la plante. Dans la nature on n'est jamais en conditions d'inhibition.
Partie 1 :
NUTRITION ET METABOLISME.
Chapitre 4 : L’ALIMENTATION EN EAU.
A L’eau du sol.
1 Liaisons de l’eau.
L’humidité est exprimée en pourcentage de la masse. C’est une notion vague, car un sable, à 10% d’eau paraît humide alors qu’une tourbe à 50% d’eau sera sèche.
L’eau peut-être libre ou liée aux constituants du sol. On trouve l’existence de plusieurs forces : osmotique, capillaire (tension superficielle), électrostatique (imbibition).
2 Potentiel hydrique et succion.
 : c’est la force d’attraction entre l’eau et le sol. Le potentiel hydrique est égal mais de signe opposé à l’énergie qu’il faut appliquer pour libérer 1 gramme d’eau. La valeur de ce potentiel est toujours inférieure à 0. Plus cette valeur est basse, plus les liaisons eau/sol sont fortes. Quand un sol se dessèche, va baisser. La succion représente la force d’attraction exercée du sol sur l’eau (cette succion est considérée comme une pression, ainsi que le potentiel hydrique).
: c’est la force d’attraction entre l’eau et le sol. Le potentiel hydrique est égal mais de signe opposé à l’énergie qu’il faut appliquer pour libérer 1 gramme d’eau. La valeur de ce potentiel est toujours inférieure à 0. Plus cette valeur est basse, plus les liaisons eau/sol sont fortes. Quand un sol se dessèche, va baisser. La succion représente la force d’attraction exercée du sol sur l’eau (cette succion est considérée comme une pression, ainsi que le potentiel hydrique).
3 La capacité de rétention.
C’est la quantité d’eau (en gramme), contenue dans 100g de sol après centrifugation à 500G ou après drainage.
4 Le point de flétrissement.
Le point de flétrissement permanent est le taux d’humidité d’un sol, pour lequel, une plante flétrie irréversiblement (sable 5%, limon 12%, argile 27%, tourbe 50%).
5 Réserve utilisable.
La réserve utilisable est la mesure de la quantité d’eau utilisable par la plante. C’est la différence entre la capacité de rétention et le point de flétrissement initial. Cette réserve représente généralement la moitié de la capacité de rétention. Un sol léger aura besoin d’un arrosage fréquent alors qu’un sol lourd n’en aura pas besoin.
B L’eau dans la plante.
1 La teneur en eau des végétaux.
Leur grande vacuole leur sert de réservoir d’eau.
Dans la plante, le xylème et le phloëme sont les vaisseaux qui conduisent les deux sèves. Le xylème est un ensemble de tissus morts, où circule la sève brute (eau +sels minéraux). Le phloëme est composé de tissus vivants où circule la sève élaborée (eau + sels minéraux + substances organiques).
 = %eau = [(MF – MS)/MF]*100 ; MS =Matière Sèche ; MF = Matière Fraîche
= %eau = [(MF – MS)/MF]*100 ; MS =Matière Sèche ; MF = Matière Fraîche
Déficit en eau : D= (m - )/m ; m = teneur maximum ; = teneur réelle.
La teneur en eau diminue avec l'âge de la plante, dans les graines, on trouve entre 5 et 10% d’eau.
2Les états et rôles de l'eau dans la plante.
On trouve l'eau sous deux états :
- L'eau libre : elle peut être en solution (dans les vacuoles ou les sèves), sous forme de vapeur (dans les méats, dans la chambre sous-stomatique).
- L'eau liée : elle peut être liée par la force osmotique, par la force capillaire (tensions superficielles), par les forces d'imbibition (force électrostatique ou colloïde).
Les colloïdes sont des macromolécules très hydrophiles, comme les argiles ou l'humus.
L'eau de constitution, est l'eau intra-moléculaire qui fait intervenir des forces très énergétiques.
Le CAH est le complexe argilo-humique.
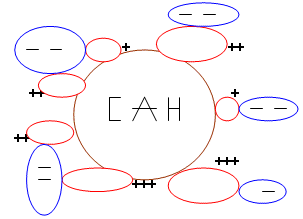
Le rôle du CAH :
- mécanique : il maintient la turgescence des cellules, donc le port des végétaux. Une perte d’eau entraîne un flétrissement.
- physiologique : dans le milieu réactionnel de la cellule, l’eau est le solvant des molécules organiques. Cette eau sert de véhicule aux aliments et permet donc la circulation des constituants de la sève.
3 la diffusion et la pression osmotique.
Le passage de molécules dissoutes à travers la membrane (avec perméabilité spécifique) entraîne un phénomène passif. De part et d'autre de la membrane, on assiste à un équilibrage grâce à l'agitation thermique : le mélange devient homogène. Avec une membrane hémiperméable, les courants d'eau vont égaliser les états énergiques des deux côtés.
La pression osmotique d'une solution, est l'attraction exercée par cette solution sur les molécules d'eau, lorsqu'elle est séparée par une membrane hémiperméable. La pression est donnée par la loi de Vant’Hoff : .v = nRT = (m/n)RT =(R.N)/(V.T)=RTC
Osmose = endosmose (ext. int.) + exosmose (int. ext.).
4 Variation de la teneur en eau dans la cellule.
L'eau est en grande partie dans la vacuole. L'eau exerce, grâce à la vacuole, au cytoplasme, au plasmodesme, une pression de turgescence sur la paroi. Les ions et molécules, dans la vacuole, lui confèrent une pression osmotique qui attire l'eau.
- La cellule dans un milieu hypotonique : l'eau rentre dans la cellule qui devient turgescente à cause d'une variation de volume cellulaire.
- La cellule dans un milieu hypertonique : l'eau sort et on assiste à une plasmolyse.
Potentiel de l'eau pression de succion ()
Potentiel hydrostatique pression de turgescence (t)
Potentiel osmotique pression osmotique (v)
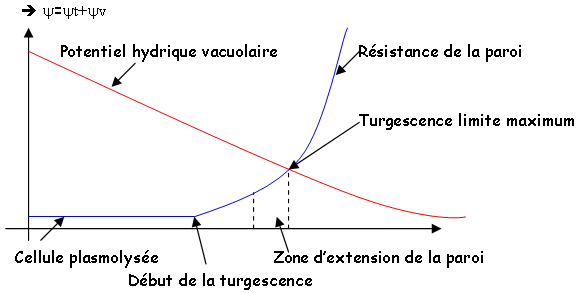
Il y a évolution =v+t à l’entrée de l’eau dans la cellule. Le xylème est lignifié pour éviter l’écrasement. On constate des variations de diamètre sur une plante pendant la journée.
5 Absorption de l’eau.
En un jour, la plante absorbe un volume d’eau équivalent à sa masse. Les plantes aquatiques absorbent l’eau sur toute leur surface. Les plantes supérieures ont leur absorption assurée par leurs poils absorbant (fixés sur les racines), et pour beaucoup d’entre elles, par des mycorhizes. C’est l’eau la moins liée au sol qui va être utilisée par la plante (on parle de liaisons osmotiques et capillaires).
6 Le transport de l'eau.
L'eau et les sels minéraux sont transportés par les poils absorbants jusqu'à l'endoderme. Voir schéma page 19. Le cheminement se fait par la loi de l'osmose (phénomènes physiques). Il y a déplacement par voie apoplastique (à travers la paroi) et symplastique (à travers les vacuoles et le cytoplasme).
Au niveau de l'endoderme, la couche cellulaire subérifiée tangentiellement (bandes de Caspary) oblige un passage par voie symplastique.
Jusqu'au cylindre central la pression (poussé) est racinaire. C’est un processus nécessitant de l’énergie métabolique.
Au niveau du cylindre central, on a un cheminement vertical qui se fait dans le xylème. Au début, la pression racinaire, devient au fur et à mesure de l'élévation par un processus physique fondé sur l'existence d'un gradient de potentiel hydrique (différence de potentiel entre le sol et l'atmosphère). L'assimilation (ou aspiration) par les feuilles permet l'évaporation (phénomène transpiratoire). L'atmosphère a un potentiel hydrique très négatif et soutire en permanence l'eau de la plante, créant ainsi un flux transpiratoire. Cette aspiration entraîne une dépression dans les vaisseaux du xylème. C'est ce phénomène qui a entraîné sa lignification.
7 Emission d'eau par la plante.
Pour qu'il y ait transpiration, il faut que soit très négatif. La différence de potentiels dépend de l'humidité relative de l'air. Le problème pour les plantes des pays équatoriaux est que l'humidité relative est de 100%. Il n'y a donc pas de différence de potentiels, donc pas de transpiration. Le transport est réalisé par les stomates aquifères hydatodes qui expulsent de l'eau. Pour cette vaporisation, il y a un besoin d'énergie solaire (95 pour cent par les stomates, 5% par la cuticule).
8 Les stomates.
Ce sont des cellules particulières situées dans les feuilles (schéma p.20). Elles ont une forme réniforme et sont accompagnées de 2 grosses cellules (cellules compagnes ou annexes) : c'est l'ostiole (là où circulent les gaz). Souvent, on observe la présence d'une chambre sous-stomatique.
Remarque : chez les monocotylédones, les stomates se trouvent sur les 2 faces des feuilles.
La structure :
- La paroi a des épaisseurs différentes : la partie interne est épaisse et cutinisée (d'où sa rigidité) ; la partie externe est fine et cellulosique (flexibilité).
- Si la cellule est turgescente, l'ostiole s'ouvre sinon elle reste fermée. C'est le mouvement de l'eau qui commande l’ouverture (K+, Cl-, malate2-).
- La mesure du potentiel membranaire met en évidence sur le plasmodesme, des pompes à protons de type ATPase qui interviennent en contrôlant la polarisation membranaire par l'expulsion de protons (besoin d'énergie).
On a mis en évidence des canaux ioniques sur la membrane, qui permettent aux ions de passer. Il l'énergie de la photosynthèse sert à faire sortir l'eau.
Remarque : les stomates à son ont les seules cellules épidermiques à posséder de la chlorophylle.
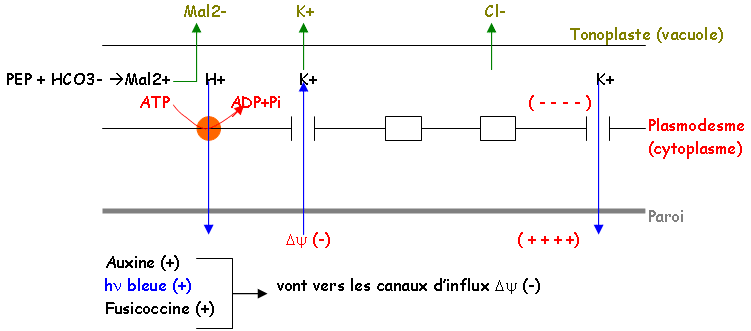
L’acide abscissique (-) (hormone) va vers les canaux d’efflux (+).
Souvent, les stomates son ont ouvert le jour (sauf chez les CAM). De plus, une faible concentration en CO2 dans la chambre sous-stomatique entraîne l'ouverture de l'ostiole.
9 Circulation des sèves.
Cette différence entre la partie aérienne et le sol fait monter la sève brute. La sève élaborée est chargée de molécules organiques (dans le liber). Cette sève est transportée bidirectionnellement, du bas vers le haut et inversement, pour atteindre toutes les parties de la plante. Cette circulation se fait avec une pression positive : on parle de flux de pression. Cette sève contient de 50 à 300 g de matière sèche par litre.
La vitesse de flux.
La vitesse de la sève élaborée est inférieure au flux transpiratoire (de la sève brute). Malgré cela, le flux de sève élaborée ne se fait pas par simple diffusion. Actuellement, ce mécanisme de circulation de la sève élaborée est mal connu. La théorie actuelle est celle ne demande de l'énergie : flux volumique sous pression (ou flux de masse) : c'est la théorie de Münch. Le chargement des tubes criblés demande de l'énergie : ce sont des potentiels négatifs.
=p++h = P-( .g.z). h : C’est le potentiel gravitationnel, il dépend de la hauteur où l'on se trouve. Voir le document page 19. Au niveau du sol, on a h=0.
.g.z). h : C’est le potentiel gravitationnel, il dépend de la hauteur où l'on se trouve. Voir le document page 19. Au niveau du sol, on a h=0.
Le phloëme : la sève élaborée s'est déchargée des solutés ( diminue). Il y a attirance du liber vers le xylème. La sève brute se charge en eau. Au sommet, La différence de pression entraîne le passage du haut du xylème vers le liber.
Partie 1 :
NUTRITION ET METABOLISME.
Chapitre 5 : LA NUTRITION MINERALE.
A Les ions minéraux, nature et importance.
Les éléments macronutritifs : K (2-4), Ca (1-2), Mg (0,1-0,7), S (0,1-0,6), P (0,1-0,5), Na, Cl, Si et Fe.
Les oligo-éléments : Mn, Zn, Ci, B, Mo, Co. On trouve ces éléments au niveau des enzymes avec différentes variations selon les espèces. On trouve le soufre chez les crucifères, le potassium chez les algues, le silicium chez des graminées, les prêles et les fougères. On trouve des variations selon les organes d'une plante. La graine est plus riche en phosphore pauvre en potassium que la plante. Les parties âgées sont plus riches en calcium alors que les parties jeunes sont riches en potassium, phosphore et azote.
B L’origine des minéraux.
Dans le sol, les ions sont en solution, ils varient selon la nature et le pH du sol. Ils sont, soit à l'état fixé dans le complexe argilo-humique (dans des solutions colloïdales), dans le complexe anionique. Si c'est une fixation paroi un cation, et si celui-ci est divalent, il fixera l'anion au sol. Si le sol a beaucoup de complexes argilo-humiques, il sera riche. La majorité des ions sont transformés par la plante pour pouvoir être utilisés. On a par exemple, les sulfates réduits au niveau des chloroplastes. Ces ions seront souvent à faibles concentrations. On trouve aussi dans la plante, des ions à concentrations élevées. On observe, chez une plante, une sélectivité pour l'absorption des ions et pour le maintien de la concentration de ces ions.
Dans le complexe argilo-humique, Les ions sont fortement liés : il y a compétition entre la plante et le complexe ionique pour capter des ions, d'où une dépense d’énergie par le végétal.
C Modalités et mécanismes de l’absorption.
1 Modalités.
Une fois l'absorption effectuée par les poils absorbants ou les mycorhizes, l'absorption est réalisée dans les cultures de manière spécifique. Par exemple, dans un verger, on fait absorber à une plante des oligo-éléments, l'absorption foliaire de fer par une plante chlorosée est le seul moyen de faire absorber ce fer sur les sols calcaires. Les ions circulent par voie symplastique (dans le cytoplasme) jusqu'au xylème puis ils passent dans la sève brute. Certains ions sont retenus au niveau des parois, comme le calcium, d'autres s'accumulent dans la vacuole sont excrétés. La vacuole est une poubelle ou une réserve.
2 Mécanismes de l’absorption.
L'absorption est sensible à la température et aux inhibiteurs métaboliques ; par exemple, une cellule morte n'absorbe pas. Ils existent plusieurs composantes dans le phénomène de transport des ions et des petites molécules. On trouve trois possibilités de pénétration : la diffusion, le transport passif (diffusion facilitée), le transport actif.
a La simple diffusion.
La membrane cellulaire permet à l'eau et aux molécules non polaires de pénétrer par simple diffusion mais aussi a quelques petites molécules polaires comme l'urée, le glycérol, le CO2, peuvent pénétrer. Ce phénomène de diffusion est exprimé par la loi de Fick.
(DQ/dT)=k.a.c ; k est le coefficient de diffusion, a, la surface de diffusion, c, la variation de concentration.
La diffusion tend vers un état d'équilibre pour que le gradient de concentration soit nul.
Le transport de petites molécules se fait grâce à des protéines membranaires de deux types. On trouve des protéines porteuses et des canaux protéiques.
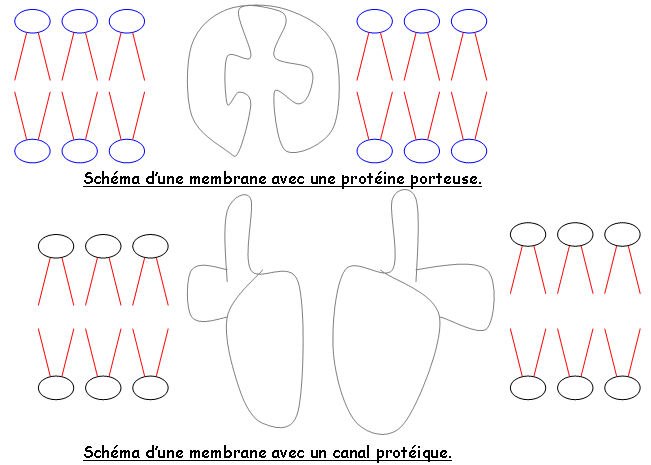
b Le transport passif et la diffusion facilitée.
Le transport se fait par des canaux protéiques et par des protéines porteuses. Si la molécule est non chargée, le transport est déterminé par le gradient de concentration. Si cette molécule est chargée, le transport est déterminé par le gradient de concentration et par le gradient électrochimique. On a donc un transport dans le sens du gradient qui entraîne un potentiel de membrane.
Le potentiel électrochimique :
µ=nF.E(2-1)+RTLog(c1/c2) (T)
=nF.E(2-1)+(RT/F)Log (c1/c2) (V)
µ=E-2,3(RT/F) pH
=E-0,06pH
Dans la diffusion facilitée le transport est réalisé dans le sens du gradient.
Le transport des molécules est réalisé par des canaux protéiques, par des protéine porteuses et par perméabilité membranaire. On trouve deux types de protéines porteuses, des protéines qui permettent un transport passif et des protéines qui réalisent ce transport grâce à de l'énergie : c'est un transport actif.
c Mécanisme actif.
Lapicque a mis en évidence le phénomène d'épictèse : c'est une absorption active des ions et des petites molécules.
Une cellule végétale dans une solution hypertonique, concentrée en saccharose est plasmolysée. Au bout d'un certain temps, la cellule redevienne turgescente : la cellule rétablie son hypertonie en absorbant des ions (ou des petites molécules) contre le gradient du potentiel électro-chimique. Ce phénomène explique qu'une cellule est capable de concentrer des ions. Ces déplacements nécessitent de l'énergie d'origine chimique, par exemple l'ATP, d'origine physique : le gradient ionique, due aux déplacements d'électrons. Cette énergie permet le fonctionnement des pompes ioniques donc le type le plus représenté est le type des pompes à protons. On trouve deux types de pompes :
- Des pompes rédox donc la circulation est réalisée grâce à des déplacements d'électrons. Ces pompes produisent de l'ATP.
- Des pompes de type ATPase : elles expulsent les protons au niveau de la membrane du plasmalème ou du tonoplasme (transports actifs). Elles utilisent de l'énergie. Le transport des protons par ces ATPases est un transport actif primaire.
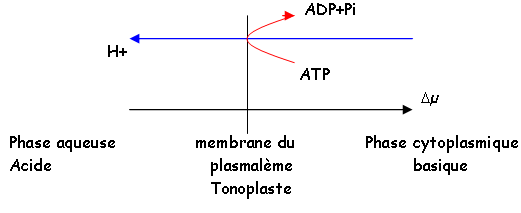
Cette émission de protons crée la «force motrice protonique », qui permet à son tour d’énergiser le déplacement d'autres espèces ioniques. On parle de transports actifs secondaires.
Ce transport actif secondaire se fait dans le sens opposé au gradient, en demandant donc de l'énergie.
S'il y a transport d'un seul soluté, on parle de système uniport.
Si deux solutés traversent dans le même sens, c'est un système symport.
Si les 2 solutés traversent dans un sens différent, c'est un transport antiport.
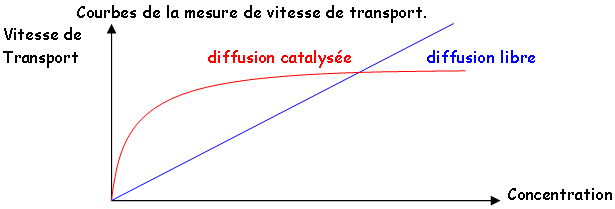
D Rôles des ions dans la plante.
1 Rôles physiques.
Les phosphates favorisent l'entrée du magnésium, alors que le calcium freine son entrée. Les ions permettent le maintien de la turgescence, du pH (système tampon), la création de potentiels membranaire qui agissent sur la perméabilité de la membrane.
2 Rôles physiologiques.
Les rôles constitutifs sont tenus par les éléments phosphorylés, comme les phospholipides, les composés phosphorylés, les nucléotides, et les acides nucléiques.
On trouve le soufre dans les acides aminés et dans les protéines.
On trouve le calcium dans les parois où ils forment avec les peptides, des pectates ; dans la vacuole il est sous forme de cristaux d’oxalate de calcium ; dans le cytoplasme il est associé à la calmoduline.
On trouve le fer (Fe) au niveau des hèmes et des cytochromes.
Le calcium se trouve dans les chloroplastes (en formant les plastocyanines) et dans les mitochondries où ils forment les cytochromes oxydases.
On trouve du molybdène dans les nitrate-réductases et dans les nitrogénases.
Le phosphore est en importante quantité pendant la floraison et dans les graines.
Le potassium est impliqué dans le métabolisme des glucides.
Le calcium se trouve le plus souvent dans les lieux de stockage des produits toxiques (en général se sont les vacuoles).
3 Quelques particularités.
a Le calcium.E
Face à la présence du calcium, on trouve deux types de plantes :
- Les calcicoles, qui tolèrent (ou supportent) le calcium. Quand la concentration en calcium va augmenter, le pH va aussi augmenter (solution basique).
- Les calcifuges, qui ne supportent pas le calcium. Leur concentration en Ca diminue en entraînant une diminution du pH (acidification).
Les plantes calcicoles peuvent modifier la composition de leur membrane pour limiter l’entrée de Ca.
b Le fer.
Sur un sol basique, le fer ne peut pas être absorbé car il précipite.
c Le sodium.
On trouve des plantes halofuges (qui ne supportent pas le sel) et des plantes halophiles (les halophytes) qui supportent le sel. Ces halophytes poussent dans les eaux saumâtres ou près de la mer (des eaux salées). Soit, elles excluent le sel, soit, elles l’accumulent dans leurs vacuoles ou dans des glandes à sel. Ces plantes ont l’aspect de plantes grasses.
Partie 2 :
DEVELOPPEMENT.
Chapitre 1 : CROISSANCE.
Le développement (en physiologie végétale) étudie toutes les modifications qualitatives et quantitatives chez une plante (de la fécondation à la mort).
Les modifications quantitatives représente la croissance (les modifications irréversibles se produisant au cours du temps). On a, par exemple, l’augmentation de taille, de volume, de masse.
On parle de différenciation quand la part prise par les modifications qualitatives vont prédominer : c’est l’acquisition de propriétés morphologiques et fonctionnelles.
A Sites et formes de croissance.
Chaque cellule va passer par une série d’étapes qui correspondent à une suite d’augmentations spectaculaires des dimensions de celle-ci.
On observe différentes étapes :
• La mérèse : c’est l’augmentation de la masse protoplastique. La mérèse est essentiellement réalisée par multiplication cellulaire (au niveau des méristèmes primaires).
• L’auxèse : c’est l’augmentation qui résulte du grandissement cellulaire (au niveau des méristèmes secondaires).
1 Au niveau de la plante et des organes.
Grâce aux méristèmes, la croissance d’une plante est en générale indéfinie (notion de taille adulte pour des organes). Une plante est soumise à deux types de croissance :
• La croissance primaire : c’est l’élongation. Elle a lieu au niveau des méristèmes apicaux (organogènes). Ce type de développement est remarquable chez tous les végétaux : c’est le port herbacé des plantes.
• La croissance secondaire : c’est l’augmentation en épaisseur. Elle a lieu au niveau des cambiums ou de zones génératrices (histogènes). Ce développement n’a lieu que chez les plantes ligneuses.
La croissance d’une plante présente des caractères commutatifs et itératifs (qui se répètent plusieurs fois).
Une plante a un développement indéfini, mais la capacité d’extension des organes est éphémère et leur grandissement se produit selon des gradients plus ou moins nets et diversement orientés suivant les organes et les espèces.
• La racine : l’élongation est réalisée par les méristèmes primaires (zone de croissance) qui permettent l’avancée dans le sol. Cette croissance (primaire) est localisée et polarisée. L’élargissement (croissance secondaire) se produit très en arrière de la coiffe.
• La tige : on ne trouve pas d’axe continu, mais des unités successives (les primarium + les ébauches foliaires). Ces unités permettent l’élongation simultanée sur plusieurs entre-nœuds successifs. Au niveau de la tige, on a un étagement du gradient de croissance qui est due à la persistance de cellules méristématiques résiduelles, juste au-dessus de chaque entre-nœuds.
• Les feuilles : l’augmentation est bidirectionnelle. L’accroissement en épaisseur est très réduit par rapport à la surface foliaire.
• Les fruits : c’est le résultat d’une hypertrophie due, dans le cas de la pomme de terre, à la croissance primaire, ou, a la croissance secondaire dans le cas du radis.
2 Au niveau cellulaire.
L’extension symplastique est effectuée avec interposition constante de cellules isodiamétriques (isotropes) et cylindriques (anisotropes).
L’extension apicale est intrusive ou extrusive.
L’extension symplastique est constante : les cellules augmentent comme un ensemble solidaire, en maintenant leurs liaisons et leurs communications.
Le cas des extensions extrusives et intrusives : la cellule acquière une autonomie plus ou moins importante par rapport aux cellules voisines.
• Extension extrusive : les cellules épidermiques (ou du rhizoderme) vont donner des poils (ex : les fibres du coton).
• Extension intrusive : elle se déroule vers l’intérieur des organes, au niveau de la lamelle moyenne (ex : les fibres de lin). L’augmentation du nombre d’individus (cellules) entraîne une augmentation des dimensions (surface, masse, …) d’un composé particulier. Pour la majorité des végétaux, on observe une augmentation de quelques centimètres par jour, avec toutefois, quelques exceptions : les asperges, 30cm/j ; les bambous, 60cm/j ; les champignons, 5mm/min.
La vitesse de croissance est donnée par la formule : v=(dl/dT) (l est un paramètre choisi).
Le taux de croissance est donné par la formule : R=V/Lo (Lo représente les dimensions initiales).
La courbe de croissance : son allure est sigmoïde. Cette courbe traduit une évolution de la plante. On peut observer quatre phases distinctes :
1. La phase de latence.
2. La phase accélérée (ou phase exponentielle) : L=Lo.e^(RT). V est proportionnelle à L, R=constante.
3. La phase linéaire : V est constante. Cette phase est parfois virtuelle, importante, ponctuelle.
4. La phase de ralentissement : c’est une phase de sénescence.
B Cinétique de croissance et variation dans la croissance.
La cinétique de croissance de la plante varie dans le temps à cause de différents facteurs : température, éclairement, humidité... La périodicité de la cinétique est dépendante de facteurs externes.
Les facteurs endogènes : par exemple, on trouve les inhibiteurs de croissance qui sont responsables de l’état de dormance d’une plante (à l’état de semence).
La variation dans l’espace : elle est due à des facteurs endogènes reliés à l’inhibition par corrélation.
Exemple : une partie d’une plante influence la croissance d’une autre partie de la même plante. Cette domination peut être due à la dominance apicale.
Partie 2 :
DEVELOPPEMENT.
Chapitre 2 : LES MEDIATEURS MOLECULAIRES DU DEVELOPPEMENT.
I Les phytohormones.
Les phytohormones sont des substances chimiques synthétisées par le végétale. Elles sont oligodynamiques : actives à faible dose. Elles agissent parfois à distance du lieu de synthèse : elles ont une influence sur le fonctionnement.
A Les auxines.
1 Nature et répartition dans la plante.
Les auxines sont synthétisées dans l’apex des tiges et des jeunes feuilles. Elles sont libres ou liées à d’autres molécules. Leur transport par le phloëme est polarisé : c’est un transport actif basipète (du haut vers le bas). Ces molécules sont des composés instables, sensibles à la lumière, à l’acidité, à l’oxygène.
2 Mode d’intervention des phytohormones.
Les effets des phytohormones sont moins prononcés chez les végétaux que chez les animaux (demande de réponse précise). Chez les végétaux, la transmission des informations est moins précise. On a du mal à séparer la transduction du signal et sa conséquence.
Les récepteurs aux phytohormones sont localisés, soit dans le cytoplasme (et accompagnent les hormones jusqu’au lieu d’action), soit sur/dans les membranes et ont alors un rôle de messagers secondaires.
L’Inositol-triphosphate : IP3 et le complexe Ca2+/calmoduline.
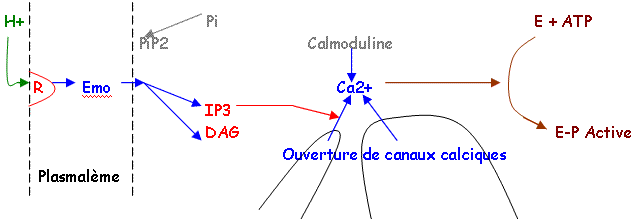
Au niveau de la membrane, les phospholipides sont transformés en PiP2. L’hormone reconnaît son récepteur et active dans la membrane, une enzyme (une lipase), qui hydrolyse le PiP2 en IP3 et DiAcylGlycérol. L’IP3 permet l’ouverture de canaux calciques au niveau de la vacuole et du réticulum plasmique : on a alors formation du complexe Ca2+/calmoduline qui va activer une kinase qui ensuite, va phosphoryler un composé «E». Ce composé "E" va assurer le transfert de l'AIA (hormone) vers le noyau.
3 Propriétés physiologiques.
Les effets sur l'élongation sont différents si l'on se trouve sur une tige ou une racine. Une dose favorable pour l'augmentation d'une tige peut-être défavorable pour l'augmentation d'une racine. En règle générale, les auxines favorisent l'accroissement des racines alors que les cytokinines favorise la croissance des tiges et des bourgeons. L'élongation fait appel à la plasticité de la paroi et à sa structure fibrillaire dans laquelle s'entrecroisent des chaînes de molécules diverses, où, entre ces chaînes, se crées des liaisons. Pour qu'il y ait élongation, il faut couper les liaisons interchaînes (intervention de l'auxine) au niveau périphérique et nucléaire.
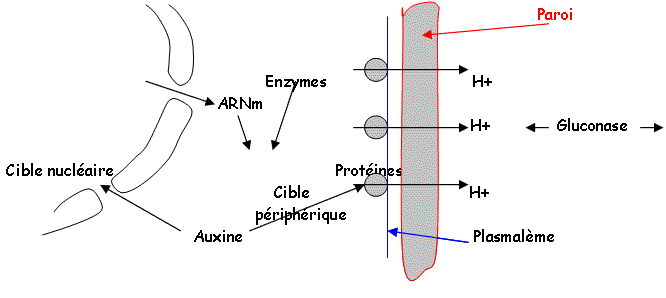
Le complexe récepteur de l'AIA dans le plasmalème, va stimuler les ATPases de celui-ci. On assiste à l'expulsion de protons vers la paroi ainsi qu'à une diminution du pH dont le but est la coupure des liaisons hydrogène.
La gluconase va couper les chaînes de polysaccharides. L'auxine va activer les pompes à protons, ce qui entraîne la possibilité de l'extension de la paroi.
La cible nucléaire : l'AIA et le récepteur soluble activent dans le noyau les gènes des enzymes responsables de la synthèse des matériaux comme les polysaccharides (pariétaux). On assiste alors à l'extension de la paroi.
En utilisant les inhibiteurs, on s'aperçoit que l'action de l'auxine se fait au niveau de la transcription.
B Les gibberellines.
1 Nature et répartition dans la plante.
Les gibberellines sont des diterpènes (molécules à 20 carbones), formées à partir de l'isoprène. Elles ont été découvertes en étudiant la maladie du ris (maladie due à un champignon).
La synthèse de ces molécules est effectuée dans des sites divers mais toujours dans des lieux de division active (à l’apex de tiges et de racines). Leur migration est polarisée. Elle se déroule dans le xylème et dans le phloëme, toujours liée à des sucres.
2 Propriétés physiologiques.
a Effets sur la croissance.
Les gibberellines permettent un allongement des entre-nœuds. Leur action se fait en synergie avec les auxines et les cytokinines.
b Autres effets.
Elles permettent la levée de la dormance des semences. Elles ont un effet antagoniste à celui de l’ABA.
c Applications.
On a trouvé des anti-gibberellines, qui sont des nanifiants : CCC, Amo16-18, phosphon : ils bloquent la synthèse des gibberellines. Les plantes ainsi traitées ont un aspect nain (au niveau des tiges mais jamais au niveau des feuilles).
C Les cytokinines.
1 Nature et répartition.
Ce sont des substances qui augmentent la division cellulaire. La première cytokinine découverte fut la zéatine, isolée dans le mais.
Ces hormones sont trouvées dans tous les tissus et sont synthétisées sur les lieux d’utilisation. On observe de fortes synthèses au niveau des racines.
2 Propriétés physiologiques.
a Au niveau cellulaire.
Les cytokinines sont présentes dans la sève brute et transportées sous forme de nucléotides de zéatine. Elles stimulent la division cellulaire en présence d’auxine : action en synergie.
b Au niveau de l’organisme.
On les trouve :
- au niveau des bourgeons, elles sont antagonistes des auxines
- pour la levée de dormance des graines,
- au moment du développement des ébauches florales,
- dans les feuilles, où elles favorisent la diminution du turn-over de la chlorophylle, en améliorant le verdissement des feuilles.
D L’acide abscissique.
1 Nature et répartition.
L’acide abscissique est un composé en C15 qui a été trouvé pendant l’abscission des fruits et des feuilles de cotonnier. Sa synthèse est réalisée au niveau des graines et des semences.
2 Propriétés physiologiques.
Il est antagoniste des gibberellines, en prolongeant la dormance des graines et des bourgeons. Il intervient aussi dans la fermeture des stomates. Il favorise la maturation des graines en stimulant l’accroissement de l’embryon et l’augmentation des réserves tout en inhibant la germination.
E L’éthylène.
On dose ce qui est dégagé par la plante. Il se forme à partir de méthionine. Il est antagoniste aux gibberellines et à un rôle dans le mûrissement des fruits.
Il inhibe l’élongation en diminuant les tiges et les racines : c’est une hormone de stress, de défense.
II Le phytochrome.
A Nature.
1 Mise en évidence
Chez les végétaux, on trouve des molécules différentes de la chlorophylle, qui sont capables de capter la lumière et d’induire sa transduction pendant le développement des végétaux. L’accroissement de la plante est modelé par la lumière : c’est la photomorphogenèse (ensemble des réactions végétales réalisées grâce à la lumière).
S’il y a absorption d’une lumière monochromatique rouge à faible énergie : le rouge clair déclenche une réponse physiologique qui est annulée par le proche infrarouge (rouge sombre). C’est la qualité du dernier éclairement qui prédomine.
Il existe dans la plante, un pigment qui, à l’obscurité, est capable d’absorber la lumière rouge. Ce pigment se transforme en une forme qui, à son tour, absorbera le rouge sombre.
La forme capable d’absorber le rouge sombre déclenche la réponse physiologique (croissance des tiges et des feuilles et arrêt de la floraison). Ces formes de pigments sont photo-convertibles.
2 Constitution.
Chromophore + protéines chaîne tétrapyrolique absorption de lumière par modifications de la position de certains hydrogènes et doubles liaisons.
Ce phénomène est comparable à ce qui se passe dans l’œil.
La lumière blanche a le même effet que la lumière rouge en réalisant une transformation qui est relativement lente (1/10ème de seconde).
B Répartition dans la plante.
Les phytochromes sont présents en faible quantité dans le végétal. Le jour, ils sont sous forme P730 (ils absorbent à  =730nm). La nuit, ces phytochromes sont sous leur forme P660.
=730nm). La nuit, ces phytochromes sont sous leur forme P660.
La forme active est la forme P730. Pendant la nuit, elle se convertie en forme inactive en subissant un turn-over.
Ce pigment est répandu chez les végétaux supérieurs (phanérogames, bryophytes et ptérydophytes).
C Les modalités d’action.
Il semble que le phytochrome agisse sur les modalités cellulaires par un changement de configuration qui modifie les propriétés membranaires, rendant possibles le passage de métabolites, de cations, d’hormones et d’ATP.
Au niveau nucléaire, les phytochrome agissent sous l’activation ou la répression de certains gènes. Quand ils sont sous forme active, ils permettent l’activation de kinases.
D Phénomènes contrôlés par le phytochrome.
• la photomorphose positive (>0) ; exemple : activation par P730.
• la photomorphose négative (<0) ; exemple : inhibition par P730.
• la photomorphose mixte ; exemple : le photopériodisme.
• La photomorphose positive : pour les plantes qui germent à la lumière, le contrôle de la germination est réalisée par le phytochrome.
L’accroissement des feuilles est stimulé par la lumière rouge et inhibée par le rouge sombre.
L’action sur le mouvement des feuilles : de nombreuses feuilles pennées (chez les légumineuses) subissent des mouvements nyctinastiques (c’est le mouvement des feuilles qui se plient). Ce mouvement a lieu si la quantité de forme active est suffisante en fin de journée. Le mouvement est due à des variations différentielles de turgescence (cellule à la base des folioles).
La synthèse des pigments entraîne la synthèse d’enzymes : PAL (PhénylalanineAmmoniaLyse), PEPcarboxylase, Rubisco.
• la photomorphose négative : c’est l’inhibition de la croissance des tiges. Il y a synthèse de Lipoxygénase qui va permettre la production d’éthylène.
• la photomorphose mixte : elle déclenche une induction ou une répression. L’exemple type en est la floraison.
Partie 2 :
DEVELOPPEMENT.
Chapitre 3 : Les mouvements des végétaux.
Pour classer les différents mouvements des végétaux, en prend en considération : la nature, le phénomène et la cause du mouvement, la nature de la partie qui bouge et la nécessité ou non d'un phénomène de croissance.
A Tropismes.
Les mouvements des organes sont déclenchés par des stimuli externes et sont orientés et liés à la croissance.
1 Le phototropisme.
La tige a un phototropisme positif alors que la racine a son phototropisme négatif. Les ramifications sont soumises au plagiotropisme.
2 Le géotropisme.
Le géotropisme est induit par l'attraction terrestre. Il peut-être positif ou négatif.
3 Le chimiotropisme.
On trouve le chimiotropisme pendant l'assimilation des substances chimiques (hydrotropisme des racines), dans le tube pollinique pendant la maturation des graines de pollen.
4 Haptotropisme ou thigmotropisme.
On parle d’haptotropisme quand une racine contourne un obstacle. On parle de thigmotropisme quand il y a accrochage d'une vrille sur un support.
B Les tactismes.
Les tactismes sont des déplacements déclenchés par des stimuli externes, orientés, et non liés à la croissance.
1 Les cas d’organites cellulaires.
Les chloroplastes se positionnent différemment face à la lumière sous l’influence du cytochrome.
2 Les cas des organismes.
Le phototactisme : c’est le déplacement vers la lumière.
Le chimiotactisme : il a lieu chez les spermatozoïdes des fougères et des mousses.
Le thermotactisme : il a lieu chez champignons inférieurs, en fonction de la température.
L’hydrotactisme : il a lieu chez les champignons inférieurs, en fonction de l’humidité.
C Les nasties.
Les nasties sont des mouvements d’organes déclenchés par des stimuli externes, non orientés, liés ou non à la croissance.
1 Epinasties et hyponasties (liés à la croissance).
Les épinasties sont des mouvements dépendants de la forme d’un organe, fixé par sa disposition anatomique. Ce sont des mouvements rapides qui concernent surtout les fleurs et les pièces florales.
Les hyponasties sont des mouvements liés à des inégalités de croissance.
2 Les nyctinasties et photonasties.
Ce sont des phénomènes non liés à la croissance.
Les photonasties et nyctinasties sont des ouvertures et fermetures de fleurs (et parfois de feuilles) en fonction d’une périodicité journalière. Ce mécanisme est lié à une intervention du phytochrome (phytochrome).
3 Les séismonasties.
Ces mouvements de séismonastie sont déclenchés par des chocs et entraînent des fermetures rapides.
D Les nutations.
Ce sont des mouvements d’organes autonomes non liés à la croissance.
Dans les cellules, on a des mouvements de cytoplasme dus aux variations de la pression de turgescence.
Dans les cellules isolées et flagellées, on va assister à un déplacement.
Partie 2 :
DEVELOPPEMENT.
Chapitre 4 : DIFFERENCIATION ORGANOGENESE ET MORPHOGENESE.
La différenciation fait appel à des notions qualitatives. Les phénomènes quantitatifs sont dus à l’acquisition de différences dans une ou plusieurs cellules.
L’obtention d’une spécialisation entraîne la différenciation en tissus et organes. Cette différenciation est possible grâce à la transmission, pendant les mitoses successives, de toutes les potentialités de la cellule originelle (du zygote). Pendant la formation des organes et des tissus, le végétal acquiert dans l’espace, une forme et une structure particulière.
Le développement d’un végétal se fait selon les informations génétiques, mais celle-ci, est modelée par des facteurs exogènes (trophiques et environnementaux) et endogènes (les hormones).
I Le cycle de développement des spermaphytes.
C’est le passage de la graine à la graine en passant par différentes phases.
La graine est en état de vie ralentie.
La multiplication végétative donne des clones qui peuvent être utilisés en horticulture. Une pomme de terre va donner des millions de plants en quelques mois.
II Développement embryonnaire : la germination.
A Le cycle de développement des semences :E
• La graine : elle provient du développement de l’ovule après la fécondation. D’autres organes, comme les fruits indéhiscents (akènes de laitue, de tournesol, …), peuvent assurer le même rôle.
• Les caryopses des graminées : pour eux aussi, on utilise le terme de semences. Ce sont des organes dont la physiologie est identique aux graines et qui sont constitués par au moins une graine (les caryopses permettent de caractériser les gymnospermes et les angiospermes) (p.33).
Les sacs embryonnaires (dans le nucelle). Ils sont composés de huit noyaux haploïdes : l'oosphère et les noyaux polaires (ou accessoires). Ce sont ces derniers qui seront fécondés. Les noyaux accessoires vont donner un tissu à 3 N chromosomes (l'albumen).
" amende " + les téguments = graine + péricarpe = fruit.
On a plusieurs types de semences en fonction de l'état final de la digestion de l'albumen par l'embryon. On distingue trois types différents :
- la digestion du nucelle par l'albumen est incomplète : il reste du nucelle qui va former la périsphère : on obtient des graines à périsphère.
- le nucelle disparaît complètement. L'albumen digère tout : c'est une graine à albumen.
- la graine est sans albumen (l'embryon l'a digéré).
Quel que soit le type de graine, on a un embryon associé à des tissus (de réserve). Cet ensemble constitue une amende. Ses réserves sont constituées de glucides, de lipides, et de protéines, avec toutefois, un composé prédominant. On a des semences protéiques chez les légumineuses, les céréales sont riches en glucides, le lin, le colza et les noix sont riches en lipides.
B La germination des semences.
1 Définition.
À l'état de vie latente, les tissus sont déshydratés (l'humidité est inférieure à 10 pour cent), les activités métaboliques sont réduites.
Pendant la germination, on trouve l'ensemble des processus qui vont du début de vie latente à la réhydratation de la graine à la sortie de la radicule.
Au sens large et dans la pratique, on englobe dans la germination, ce qui concerne la levée de la graine (la croissance de la radicule et de la gemmule) jusqu'à l'apparition des premiers organes aériens.
2 Les phénomènes morphologiques.
On trouve deux types de germination :
- La germination épigée du haricot : la radicule va donner les racines, la tigelle va donner l'hypocotyle. La gemmule donne l'épicotyle et les feuilles. Ce sont des graines sans albumen dont les cotylédones vont dégénérer.
- La germination hypogée (c'est le cas de la sève). Il n'y a ni de tigelle ni d'hypocotyle. Ici, les cotylédones sont des organes de réserve.
Chez d'autres végétaux, les cotylédones vont donner les premières feuilles.
3 Phénomènes physiologiques et biochimiques :
La courbe théorique d’imbibition des semences.
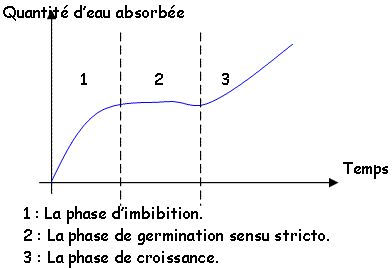
Dans la phase 1, l'eau va vers l'embryon : on assiste à la reprise des activités métaboliques (il faut de l'énergie). Dans ce cas, la respiration est très active. Chez certaines graines, l'énergie vient de la fermentation.
La seconde phase se termine par la sortie de la radicule. Cette sortie marque le passage d'un état physiologique réversible à un état irréversible.
La troisième phase : on assiste à la croissance et au développement des racines et de la tige.
Les réserves sont mobilisées dès la première phase (on a des synthèses d'hormones comme les gibberellines).
4 Conditions de germination.
a Les conditions externes (indispensables).
Les conditions externes indispensables sont, la présence d'eau, d'oxygène, et d'une température conforme aux activités métaboliques. On a aussi la lumière qui favorise (et qui est parfois indispensable à) la germination.
Chez les plantes, on trouve plusieurs types de photosensibilité :
- La photosensibilité positive : elle est présente chez 70% des semences, c'est un besoin de lumière.
- La photosensibilité négative : c'est un cas rare que l'on trouve chez les liliacées.
- pas de photosensibilité : on retrouve ce cas chez la majorité des plantes cultivées.
La sensibilité dépend du cytochrome (pour la synthèse de P730). Il peut y avoir besoin de certains sels, par exemple KNO3. La coumarine inhibe la levée des graines.
b Les conditions internes.
Une semence doit être mûre : on a donc, chez la plante, une obligation de maturité. Toutefois, chez certaines espèces, la maturité n'est pas terminée quand la graine et le fruit tombent. Cette maturité est obtenue sur le sol.
La longévité du pouvoir germinatif varie selon les espèces et peut durer quelques jours à plusieurs années. Ce pouvoir germinatif est souvent lié à la nature des réserves. Par exemple, les lipides durent peu de temps.
Quand on se trouve dans de bonnes conditions, certains organismes sont malgré tout insensible aux conditions externes : ils sont toujours en état de dormance et le retour à la vie active ne se fera qu'après une transformation externe qui restitue la sensibilité aux différents facteurs : c'est la levée de dormance.
La dormance :
- Au niveau tégumentaire : les enveloppes bloquent la germination. Etant très solides, ces enveloppes créent une résistance mécanique, une imperméabilité à l'eau, à l'oxygène et aux substances inhibitrices. La levée se fait par une altération des enveloppes (c'est une scarification).
- Au niveau embryonnaire : ce type de dormance est levé par le froid humide (c'est une dormance psychrolabille) cette dormance est sous contrôle hormonal. Au cours de la stratification, il y a une diminution de la quantité d'acide abscissique et une augmentation de la quantité de gibberellines.
III La différenciation des autres organes.
A La rhizogenèse.
La formation de nouvelles racines comporte une phase de différenciation des cellules péricycliques, face aux pôles ligneux. L'origine des racines est profonde.
B La caulogenèse.
La caulogenèse correspond au développement des bourgeons et des tiges. C'est l'anneau initial qui donne naissance aux feuilles et aux tissus périphériques. Au centre de l'anneau initial, on trouve un méristème d'attente qui se développe. À la floraison. Le méristème médulaire forme la moelle.
Dans le cas de la tige, on trouve des méristèmes intercalaires qui complètent la croissance de celle-ci. Les tiges et les feuilles ont une origine superficielle.
Le développement des organes dépend des conditions ambiantes, des potentialités du génome et du fonctionnement des autres organes : c'est la corrélation morphogénétique.
C Les corrélations morphogénétiques.
Ces corrélations peuvent être de nature différente :
- de nature trophique : il y a compétition entre les différents organes pour la distribution des substances nutritives (il y a fourniture d'un organe par un autre organe pour un composé précis).
- de type hormonal : l’auxine est synthétisée dans les bourgeons mais elle agit dans les racines.
La corrélation racine/tige : la racine alimente la tige en eau et en sels minéraux. Dans la racine, il y a synthèse de gibberellines et de cytokinines (en majorité). Chez les plantes pérennes (vivaces) et bisannuelles, la partie qui reste pendant l'hiver correspond aux racines.
La corrélation tige/racine : tous les produits de la photosynthèse et les hormones produites dans la partie supérieure vont aller alimenter les racines.
La corrélation bourgeons/bourgeons : on observe une dominance apicale : celle si est responsable de la forme des plantes et des arbres (en particulier) qui est du à l’auxine. C'est une dominance plus ou moins forte. On trouve différents types de dominances :
- la dominance apicale stricte. Elle a lieu chez les conifères.
- pas de dominance.
- la dominance par acrotonie du débourrement entraîne le port arborescent comme chez les arbustes.
- la basitonie du débourrement. Les bourgeons du bas se développent en premier : c'est le port buissonant.
IV Les capacités d’organogenèse des végétaux.
Cette capacité d'organogenèse est représentée par des techniques comme le bouturage, le marcottage et par la reproduction asexuée. On peut aussi inclure la technique de clonage. On remarque aussi la totipotence des cellules végétales : on peut obtenir une plante à partir d'une cellule isolée. On peut assister à une embryogenèse somatique à partir de tissus diploïdes ou haploïdes. Cette dernière technique permet la création de plantes transgéniques (créées à partir de cellule seule) pour obtenir un meilleur rendement.
V Le développement reproductif (physiologie de la floraison).
A Acquisition de l’aptitude à fleurir (formation d’ébauches).
1 Maturité de floraison.
Une plante ne fleurie que si elle atteint un certain développement végétatif (pour un arbre fruitier, il faut attendre cinquante à sept ans, pour un pied de tomate il faut attendre la formation de treize entre-nœuds). Chez beaucoup d'espèces, l'aptitude à fleurir ne sera acquise qu'après l'intervention de facteurs externes comme la température et la lumière. De plus, la nutrition de la plante intervient (un excès d'azote favorise le développement végétatif et inhibe la floraison). Il faut [C]/[N]=15à20. Il existe des exceptions, chez les agrumes il y a un grand besoin en azote.
2 L’action de la température..
a La vernalisation.
Les plantes monocarpiques ne fleurissent qu'une fois dans leur vie. Elles peuvent être annuelles, bisannuelles, pluriannuelles comme les agaves. Les plantes polycarpiques forment des fleurs tous les ans. La température agit généralement par le phénomène de vernalisation : il y a floraison si seulement, la plante a été exposée à basse température. Le contrôleur de l'acquisition de l'aptitude à fleurir est réalisé par un abaissement temporaire de la température. Par exemple, le blé tendre (variété d'hiver que l'on sème en autonome) germe au début de l'hiver et forme des fleurs au printemps. Si ce blé est planté au printemps, il reste à l'état végétatif. La variété de blé de printemps, si elle est semée à l'automne, va geler. Si elle est semée au printemps, elle va fleurir avec un cycle court (moins productif). Le traitement par le froid peut-être à remplacer par les hormones, par la décapitation, par de fortes températures. Par exemple, les plantes de types scrofulaires, qui ont besoin de - 3°C pendant huit semaines peuvent être traitées à 32°C pendant trois semaines.
Les autres types de réaction face à la température :
• Les plantes peuvent être indifférentes.
• Elles peuvent être préférantes : elles n'ont pas besoin du froid, mais il accélère le virage floral. C’est le cas des céréales (d'hiver), des légumineuses et des plantes bisannuelles.
• On trouve aussi des plantes qui en ont un besoin absolu de vernalisation, se sont des plantes qui passent l’hiver à l'état de rosette et dont les bourgeons doivent être vernalisés.
b Le thermopériodisme.
Le thermopériodisme est l'aptitude des plantes à fleurir, si les alternances de température sont correctes (soit saisonnières, soit quotidiennes).
- Thermopériodisme saisonnier : on le trouve chez les bulbes, les rhizomes comme le muguet, les plantes vivaces, les arbres fruitiers. La différenciation florale se fait l'été et l'hiver permet de lever la dormance.
- Thermopériodisme quotidienne : on le retrouve chez le pois ; la floraison n’est accélérée que si la température nocturne est élevée.
B La mise à fleur : le photopériodisme.
Quand l'aptitude à fleurir est acquise, la plante va être apte à la mise à fleur. Pour les plantes des régions du sud de la France, la floraison est contrôlée par la photopériodicité journalière (toutes les 24 heures). L'hémépériode correspond à la période claire du jour, alors que la myctipériode correspond à la phase sombre.
Les différents types de plantes :
- on trouve des plantes indifférentes à la lumière : il y aura des fleurs quelle que soit la durée du jour par rapport à celle de la nuit. C'est le cas du cerisier, de la tomate, du maïs...
- on trouve des plantes aphotiques qui ne forment leurs ébauches florales qu'à l'obscurité. C'est le cas de la jacinthe, de la pomme de terre.
- on a aussi des plantes de jours courts (elles sont dites myctipériodiques). Elles ne fleurissent que si l'hémipériode (eupériode) est inférieure à la période critique. La période obscure ne doit pas être interrompue. C’est le cas du chrysanthème, du topinambour, des chalanchloës.
- les plantes de jour long (plantes hémipériodiques). La période favorable est supérieure à la période critique. C'est le cas du tournesol, des céréales...
On a des plantes absolues et des plantes préférantes. Il ne faut pas oublier que la nutrition est aussi impliquée dans la capacité de mise à fleur. Le cytochrome est lui, responsable de la floraison.


