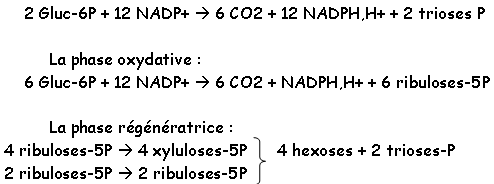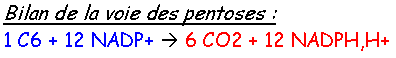
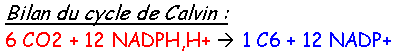
Chapitre 3 :
Les mitochondries et la respiration.
I Introduction.
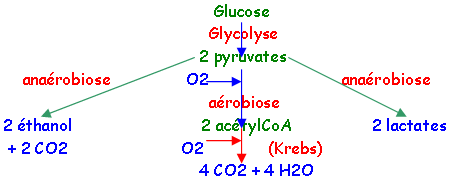
La respiration a lieu chez tous les organismes qui utilisent l’O2 comme accepteur final et le pyruvate pour donner de l’ATP, de l’eau et du CO2.
II La mitochondrie.
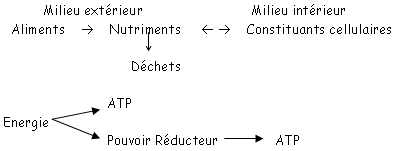
La fonction principale de la mitochondrie est l’approvisionnement de la cellule en ATP par des processus biochimiques qui sont identiques dans chaques cellules.
La mitochondrie est un organite à double membrane :
- dans la matrice se déroule la décarboxylation oxydative du pyruvate et le début du cycle de Krebs
- dans la membrane se trouve la chaîne respiratoire.
Grâce à ses crêtes (invaginations), la membrane interne est 4 ou 5 fois plus grande que la membrane externe.
· La membrane externe est perméable et reliée au réticulum endoplasmique.
· La membrane interne est imperméable, d’où la présence de transporteurs souvent spécifiques qui permettent le passage de molécules. Pyruvate, phosphate, ATP, ADP et navettes servent de transporteurs.
Dans la matrice, on note la présence :
- d’enzymes :
· pour la chaîne respiratoire
· pour la formation d’ATP
· pour la dégradation d’acides gras.
- d’ADN et de ribose.
Dans la phase aqueuse (entre les deux membranes), il n’y a pas de réaction enzymatique.
III La décarboxylation oxydative du pyruvate.
A Mécanisme.
L’entrée du pyruvate dans la mitochondrie active sa prise en charge par la pyruvate déshydrogénase (complexe enzymatique).
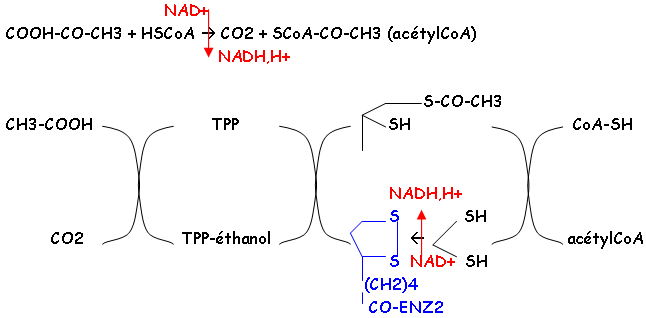
Remarques :
- ENZ = pyruvate déshydrogénase
- Coenzyme : TPP = Thyamine pyrophosphate ; l’acide lipoïque permet le transfert d’un groupement par une oxydoréduction.
B Origine et rôle de l’acétylCoA.
Il provient de la dégradation de lipides et de glucides qui va donner les glycérides qui pourront ensuite donner des acides gras.
- Chez les animaux, on a deux types d’acides aminés :
· les acides aminés cétogènes vont donner, en se dégradant, l’acétylCoA.
· les acides aminés nucléoformateurs qui peuvent participer à la synthèse de glucides.
- Chez les végétaux, l’acétylCoA vient du glycoxylate.

IV Le cycle de l’acide citrique (le cycle de Krebs).
A Les différentes réactions.
AcétylCoA + AOA 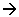 C6 (acide citrique)
C6 (acide citrique)
(C2) (C4)
La série de transformation du C6 se fait avec : + 2 CO2, + du pouvoir réducteur, + ATP.
Les enzymes situées dans :
- la matrice catalysent les réactions 1, 4, 6, 7, 8 et 10
- la membrane catalysent les réactions 2, 3, 5 et 8.
Déroulement du cycle de Krebs :
1 : la citrate synthase donne le citrate (C6) qui est symétrique.
2 et 3 : l’aconitase réalise une déshydratation puis une hydratation qui permettent un déplacement de la fonction alcool è isocitrate.
4 et 5 : l’isocitrate déshydrogénase è l’acide a-cétoglutarique (C5).
6 : l’a-cétoglutarate déshydrogénase è succinyl-CoA oxydé et décarboxylé.
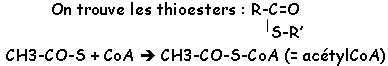
7 : libération d’énergie è succinate (C4) grâce à la succinate thiokinase. On a une libération de CoA (radical thiol).
8 : par oxydation, la succinate déshydrogénase (flavoprotéine Fe-S qui fonctionne avec du FAD) donne le fumarate.
9 : la fumarase par hydratation donne le malate.
10 : la malate déshydrogénase (par oxydation) donne l’AOA.
Un tour de cycle donne :
+ 2 CO2
+ 3 NADH,H+
+ 1 FADH2
+ 1 ATP
Au niveau du fumarate, on a un inhibiteur compétitif (le malomate : COOH-CH2-COOH) qui bloque Krebs par analogie stérique.
B Bilan.
La glycolyse donne 2 pyruvates.
2 CH3-CO-COOH 2 CH3-CO-ScoA + 2 NADH,H+ + 2 CO2
2 CH3-CO-ScoA 4 CO2 + 6 NADH,H+ + 2 FADH2 + 2 ATP (ou GTP) + 2 HSCoA.
La respiration aérobie donne de l’énergie par combustion de molécules en libérant du CO2. On trouve bien ce type de réactions dans le cycle de Krebs, ce qui montre que celui-ci fait bien parti de la respiration aérobie.
Résumé :
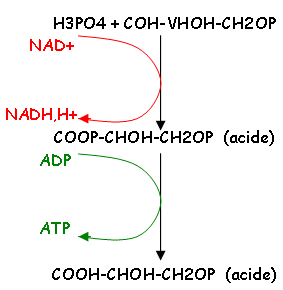
· Gluc 2 Pyruvates + 2 ATP + 2 NADH,H+ (glycolyse)
· 2 pyruvates 2 acétylCoA + 2 CO2 + 2 NADH,H+ (décarboxylation oxydative)
· 2 acétylCoA 2 FADH2 + 2 ATP + 4 CO2 + 6 NADH,H+ (Krebs)
2 FADH2 + 4 ATP + 6 CO2 + 10 NADH,H+ (Chaîne respiratoire)
10 NADH,H+ à 10 ´ 3 = 30 ATP
+ 2 FADH2 à 2 ´ 2 = 4 ATP
+ 4 ATP (Chaîne respiratoire)
38 ATP.
C Place du cycle de Krebs dans le métabolisme intermédiaire.
Krebs est une passerelle entre l’anabolisme et le catabolisme.
· AOA Aspartate
· a-cétoglutarate glutamate
· succinyl composés des porphynes
· acétylCoA
Les réactions anaplérotiques permettent la ré-alimentation du cycle de Krebs.
1 La carboxylation du pyruvate.
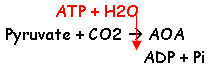
2 La carboxylation du PEP.
PEP + CO2 + GDP AOA + GTP (cette réaction fournie de l’énergie sous forme de GTP ou d’ATP).
3 Le cycle du glycoxylate.
On a synthèse de glucides à partir d’acétylCoA issu de la dégradation des lipides.
Acides gras acétylCoA cycle du glycoxylate glucides (par nucléogenèse). Succinate
Remarque : chez les plantes riches en lipides, le glycoxysome est un organe spécialisé à une seule membrane.
D La régulation du cycle de Krebs.
· La vitesse de cycle est différente selon les différents besoins en énergie.
· Les points de régulation :
- en 1 : inhibition par l’ATP
- en 4,5 : activation par l’ADP
- en 6 : inhibition par le succinyl et par le NADH,H+
- à l’entrée du pyruvate.
V Le catabolisme des acides gras.
Les glycérides (glycérol estérifié par un ou plusieurs acides gras) sont des lipides de réserves.
A L’activation des acides gras.
Ils sont hydrolysés dans le cytoplasme par des lipases. Ils donnent alors du glycérol + des acides gras puis du Dihydroxyacétone (DHAP) qui passera dans la glycolyse.
Les acides gras vont dans la mitochondrie et y sont dégradés en AcylCoA par une enzyme thiokinase (située sur la membrane externe de la mitochondrie).
R-COOH + ATP + CoA-SH + H2O RCO-S-CoA + AMP + 2 Pi
Remarque : la carnitine permet le transport des acides gras dans la matrice.
B La b-oxydation.
C’est une dégradation des acides gras.
L’oxydation se fait en C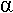 et C
et C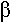 grâce à l’AcylCoA déshydrogénase qui fonctionne avec du FAD. Le produit de la réaction est l’énoylCoA (sous forme trans).
grâce à l’AcylCoA déshydrogénase qui fonctionne avec du FAD. Le produit de la réaction est l’énoylCoA (sous forme trans).
C Les cas particuliers.
· L’éthylène hydratase (ou crotonase) + H2O
· Thiolase AcylCoA(n-2) + acétylCoA. hydroxy-acylCoA.
Remarque : l’AcylCoA remonte en haut de l’hélice réactionnelle pour effectuer un autre tour. A la fin on obtient deux acétylCoA.
· Si on a un acide gras insaturé (forme cys), il y a action d’une isomérase avant l’action de l’hydratase.
· Si le nombre de carbone est impair, au dernier tour de spire, on obtient un acétylCoA et un propanoylCoA (C3). Ce dernier sera décarboxylé, donnera alors de l’acide succinique qui ira à son tour alimenter le cycle de Krebs.
D Bilan.
Un acide gras à nombre pair de carbone :
- à chaque tour, on obtient : + 1 FADH2 + 1 acétylCoA + 1 NADH,H+
- au dernier tour, on a : + 1 FADH2 + 2 acétylCoA + 1 NADH,H+
VI La chaîne respiratoire : transfert des électrons et phosphorylation oxydative.
A Les constituants de la chaîne respiratoire.
La membrane interne de la mitochondrie.
Elle est le lieu de la formation de l’ATP et du pouvoir réducteur. Ceux-ci sont synthétisés grâce aux processus énergétiques réalisés par une de transporteurs de la chaîne oxydative. Les transferts d’électrons ont comme accepteur final l’O2. Ce transfert d’électrons est possible grâce à la différence de potentiel rédox. Si cette différence de potentiel est suffisamment grande, une phosphorylation oxydative permet alors la synthèse d’ATP.
L’évolution de la chaîne à partir des potentiels.
Dans la mitochondrie, en présence d’O2 et à l’équilibre, les transporteurs de l’extrémité la plus réductrice sont sous forme réduite, alors que ceux qui sont proches de l’O2 sont sous forme oxydée.
A partir de la membrane de la mitochondrie, on isole surtout des complexes multi-enzymatiques.
1 Les flavoprotéines.
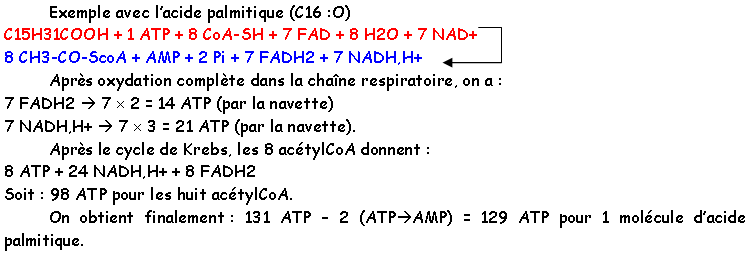
a La NADH déshydrogénase ou FP1 ou complexe 1.
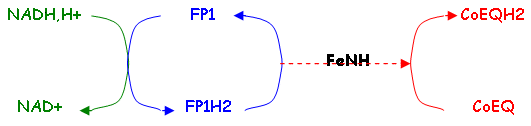
FeNH est une protéine à fer non hémique.
La NADH déshydrogénase est une flavoprotéine dont le poids moléculaire est 150.000KDa. Cette enzyme va oxyder le NADH,H+.
b La succinate déshydrogénase ou FP2 ou Complexe 2.
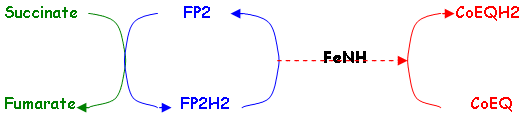
FP2 est liée à la membrane et oxyde le succinate en fumarate ; son poids moléculaire est de 250.000 KDa.
c L’AcylCoA déshydrogénase ou FP3.
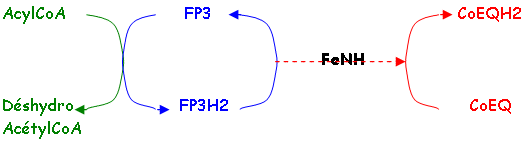
d La glycérol-phosphate déshydrogénase ou FP4.
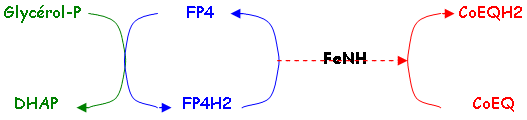
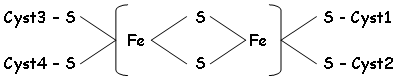
2 Les protéines à fer non hémique (FeNH) ou protéines Fer-Soufre (F-S).
Ces protéines servent d’intermédiaires entre le CoEQ et le coenzyme des protéines.
3 Les coenzymes quinoniques CoEQ ou ubiquinoniques.
Ce sont les transporteurs liposolubles de la membrane. De plus, ceux-ci fonctionnent seuls.
4 Les cytochromes.
Ce sont des hémoprotéines dont le noyau tétrapyrolique comporte du Fe2+ ou du Fe3+.
On a plusieurs types de cytochromes qui sont différenciés selon les substituants du noyau, la partie protéique et le nombre de liaisons impliquant le fer (le fer présente six valences de coordination possibles). Quatre des liaisons du fer se font avec le noyau, les deux autres peuvent se faire avec la partie protéique. Quand les six valences sont bloquées, elles donnent de simples transporteurs d’électrons.
· Les cytochromes a + a3 ou cytochromes oxydases :
Ce sont les éléments ultimes de la chaîne, ils sont insolubles et contiennent du cuivre. Ils possèdent un hème de type A. Ce dernier possède une valence libre qui est responsable des inhibitions possibles par le cyanure ou le CO. Ce sont eux qui amènent les électrons au bout de la chaîne.
4 électrons + O2 + 4 H+ 2 H2O.
· Les cytochromes c :
Ils sont solubles, liés à leur protéine par des liaisons thioesters et de coordination.
· Les cytochromes b et c1 :
Ils sont fortement liés à la membrane par des liaisons de même type que celles des cytochromes de type c. Seule la liaison à la protéine change.
B Les transferts d’électrons.
En mesurant le potentiel rédox et en utilisant des inhibiteurs, on a pu définir l’ordre de déroulement des différentes étapes.
Toutes les étapes qui arrivent au CoEQ assurent le transfert de protons et d’électrons. Au niveau du CoEQ, les électrons se séparent des protons et vont migrer le long de la chaîne respiratoire à la face interne de la membrane interne.
Les électrons se lient aux protons qui migrent vers la face externe de la membrane interne (vers la phase aqueuse).
Remarque : Le cyanure bloque la respiration.
C La phosphorylation oxydative.
1 Aspects thermodynamiques.
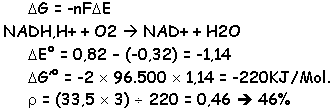
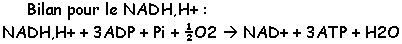
L’oxydation par phosphorylase fournie en ATP et le reste de la production est dissipé sous forme de chaleur.
2 Couplage d’énergie et phosphorylation
On va mesurer la quantité de Pi incorporé et la quantité d’O2 consommé.
· P/O = 3 quand le NADH,H+ est oxydé.
· P/O = 2 quand le FADH2 est oxydé (quand [S] est le succinate).
La phosphorylation contrôle le transfert d’électrons. L’ADP est un régulateur de la r régulation (de la respiration).
Le dinitrophénol et le dicoumarol bloquent la phosphorylation tout en permettant le transfert des électrons : on a donc une modification de la perméabilité de la membrane.
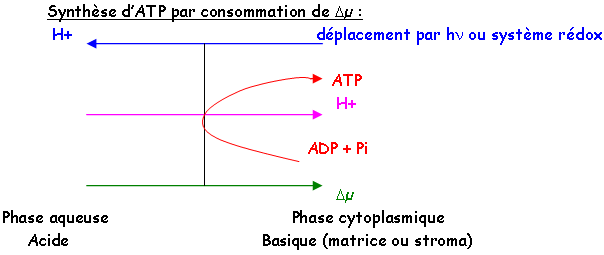
3 Mécanisme de couplage : théorie chimio-osmotique de Mitchell.
C’est la théorie qui s’accorde le mieux avec les résultats expérimentaux.
Une partie de l’énergie libérée pendant le transfert d’électrons est captée par les éléments de la chaîne respiratoire afin d’expulser les protons.
Ce mouvement va créer :
- Un gradient de pH : [H+] de la matrice < [H+] extérieure matrice è pH matrice > pH extérieur.
- Un gradient de potentiel : c’est le potentiel de membrane.

Le gradient est annulé par le retour des protons vers la matrice. Retour qui est contrôlé par l’ATP synthase.
Le retour d’un proton entraîne une énergie de –21,6KJ/Mol, alors que la synthèse d’un ATP demande –33,5KJ/Mol pour la synthèse d’un ATP, l’ATP synthase devra fournir une énergie telle que  µ
µ  G. Il faut donc que 2 H+ soient transportés pour qu’il y ait synthèse d’un ATP.
G. Il faut donc que 2 H+ soient transportés pour qu’il y ait synthèse d’un ATP.
L’orientation des ATPases membranaires :
1 : Le gradient électrochimique de H+ est toujours orienté de la phase aqueuse acide à la phase cytoplasmique basique.
2 : La synthèse ou l’hydrolyse de l’ATP a toujours lieu sur la face plasmique de la membrane.
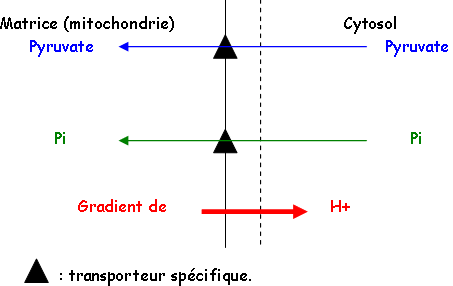
D Transports des métabolites dans la mitochondrie.
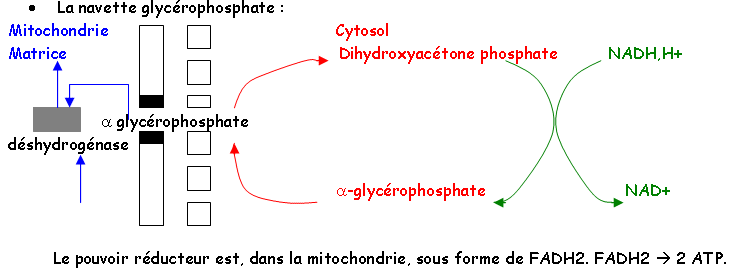
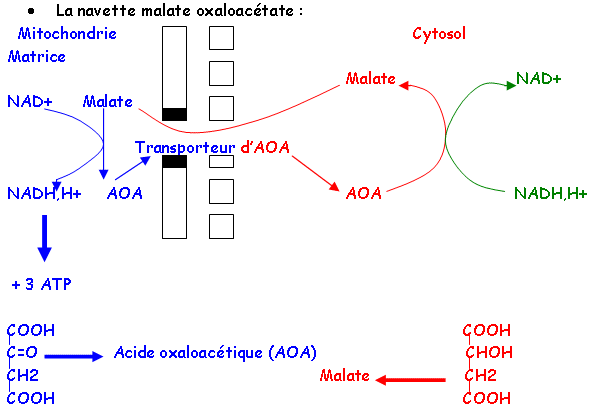
Les échanges permanents entre le cytoplasme et les mitochondries permettent l’entrée de pyruvate, de pouvoir réducteur d’ADP et de Pi grâce à des navettes de deux types : glycérophosphate et oxaloacétate.
Conclusion et bilan énergétique :
Glycolyse :
1 Gluc 2 acides pyruviques + 2 ATP + 2 NADH,H+ +
Décarboxylation oxydative :
2 acides pyruviques 2 acétylCoA + 2 NADH,H+.

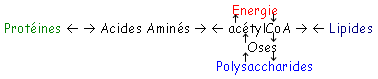
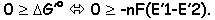

 2 trioses phosphates (aldolase)
2 trioses phosphates (aldolase)