Biologie Cellulaire
Rappels.
I Les tissus épithéliaux.
Les tissus épithéliaux sont composés de cellules jointives. Ces cellules vont établir une barrière entre deux milieux de nature différente. Les jonctions réunissent les cellules pour former un épithélium réglant la cohésion entre cellules et les échanges entre celles-ci.
A Les jonctions assurant la cohérence de l’épithélium.
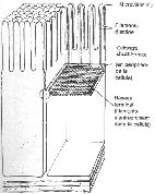
La jonction la plus importante est la jonction fermée (ou tight junction) ; elle définie une polarité : le pôle apical est situé entre deux jonctions fermées ; à l’opposé, on trouve le pôle basal.
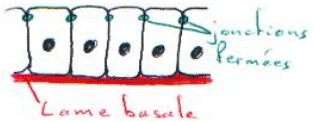
Le pôle apical est tourné vers le milieu extérieur alors que le pôle basal est orienté vers la lame basale (où il y a production de cellules épithéliales). Le pôle basal est tourné vers le circuit de capillaires sanguins qui est souvent placé à la base de la lame basale.
Il existe dans la membrane plasmique de ces cellules, des protéines (les occludines) qui ont la propriété de traverser cette membrane et d’interagir avec les occludines de la cellule voisine : il y a établissement d’une barrière entre les deux membranes plasmiques par l’interaction du domaine extracellulaire de ces protéines (interaction par les ions Ca2+).
Pour séparer les cellules, il faudra utiliser de l’EDTA qui permet la destruction de ces liaisons.
Par leur domaine cytosolique, les protéines sont associées indirectement avec les éléments du cytosquelette (les microfilaments) : il y a immobilisation de ces protéines à l’apex des cellules.
On peut aussi voir la position des protéines intégrées dans la membrane, organisées de façon particulière (en lignes anastomosées) entre elles, sur plusieurs rangées. On a un réseau de lignes anastomosées autour de la cellule : c’est la ceinture continue.
Si l’on place dans un des milieux, par exemple extracellulaire, un marqueur opaque aux électrons, on constate qu’il n’y a pas de passage. Si le marqueur se trouve dans le sang, il ne diffuse pas jusque dans le milieu extérieur. à S’il y a un passage, il est géré par les cellules.
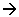 L’épithélium est une véritable barrière entre deux milieux.
L’épithélium est une véritable barrière entre deux milieux.
Les protéines forment des lignes anastomosées. Elles sont adhérentes l’une à l’autre et organisées en ligne.
Au-dessous de la jonction fermée, on trouve un autre dispositif comparable : la ceinture d’adhérence. Cette ceinture fait le tour de la cellule.
Cette ceinture est en relation avec les microfilaments d’actine. Dans le cas des entérocytes, les filaments d’actine s’organisent dans le cytoplasme en formant un plan : le réseau terminal.
Les desmosomes.
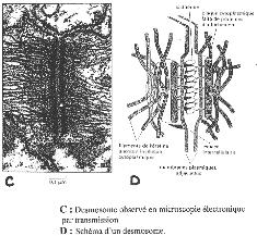
Les desmosomes sont des formations d’un micromètre de diamètre. L’espace intercellulaire est dilaté et dans la région intracellulaire sous membranaire, sont différenciées des plaques opaques aux électrons. Dans la membrane, au niveau de ces desmosomes, il existe des protéines intégrées (cadhérines) qui interagissent entre elles d’une cellule à l’autre.
Par leur domaine cytosolique, ces cadhérines agissent sur les protéines de la plaque (les desmoplaquines). Elles sont reliées avec des filaments intermédiaires (tonofilaments à kératine). Ces derniers filaments, dans une cellule épithéliale, s’organisent en allant d’un desmosome à l’autre : ils donnent une tonicité à la cellule. Cela équivaut au squelette de la cellule.
Sur la face basale, il existe des jonctions augmentant l’adhérence mais qui ont pour partenaire la lame basale et non une autre cellule. On trouve surtout deux types de jonctions :
- Les hémidesmosomes : Ce sont des formations asymétriques ; l’implantation des cadhérines se fait dans la lame basale.
- Les contacts focaux : Ce sont des différenciations locales qui permettent un ancrage transitoire à un support (normalement la lame basale). Cette différenciation transitoire est labile et permet à la cellule de se positionner sur un support et d’y migrer. La jonction avec la membrane basale est réalisée par des intégrines (protéines) qui traversent la membrane plasmique. Leur domaine extracellulaire interagit surtout avec les protéines de la lame basale (particulièrement la fibronectine). Par le domaine intracellulaire, il y a interaction indirecte avec les microfilaments d’actine.
B Les jonctions permettant la communication entre les cellules épithéliales.
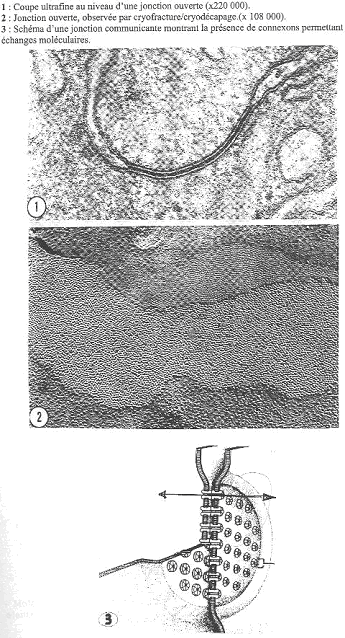
Ces jonctions sont sur les faces latérales ; leur but est de transférer des informations d’une cellule à l’autre. è L’épithélium est une unité fonctionnelle.
On peut observer une coupe transversale avec une jonction ouverte qui occupe un espace délimité de la membrane
On peut observer une densité importante de protéines au niveau de la jonction. Ces protéines sont des connexines ; elles sont organisées les unes par rapport aux autres par groupe de 6 et forment un connexon. Un connexon est un cylindre au centre duquel les molécules solubles dans l’eau pourront diffuser d’une cellule à l’autre. Un canal d’une cellule possède un canal en vis-à-vis dans la cellule voisine. Des petites molécules d’une taille inférieure ou égale à 1200Da (»1 nucléotide) peuvent passer par ce canal. è Une information apportée à une cellule peut ainsi facilement diffuser aux cellules voisines.
Cette jonction est dynamique : elle peut s’ouvrir et se fermer. Si la quantité de Ca augmente, il va y avoir fermeture du connexon. Si un trauma a lieu dans un épithélium, il va y avoir fermeture des jonctions ouvertes.
L’épithélium fonctionne comme une entité répondant à des variations du milieu extérieur
On peut observer des épithéliums différents selon des critères fonctionnels :
- Epithélium à cellules endocrines ; facilement identifiable car les cellules sont capables de synthétiser des protéines et de les exporter vers le pôle basal.
- Une cellule exocrine, elle, exportera ces produits de synthèse vers l’apex (milieu extérieur).
- La cellule absorbante : si une cellule capte des molécules au niveau de son pôle apicale et les fait transiter vers le pôle basal, c’est une cellule absorbante ; à l’inverse, une cellule excrétrice fera passer les molécules du milieu intérieur vers le milieu extérieur.
Les épithéliums peuvent être classés en fonction de caractères morphologiques. Le premier critère est le nombre d’assises composant l’épithélium.
- Une assise cellulaire : Epithélium simple. Il faut ensuite regarder la forme générale des cellules.
· Si elles exposent vers la base la plus grande surface de contact et ont une faible hauteur : c’est un épithélium squameux.
· Si la surface de la base est à peu près aussi grande que la hauteur, c’est un épithélium cuboïdal.
· Si les cellules sont étroites et hautes, c’est un épithélium palissadique.
- Plusieurs assises cellulaires. Il faut regarder l’assise supérieure de cellules (les cellules proches de la lame basale sont souvent cuboïdales).
· On peut avoir, selon les cellules de surface, un épithélium stratifié squameux, cuboïdal, palissadique.
· Selon la différenciation des cellules de surface, on parlera d’épithélium strié si l’on est en présence de microvillosités ou d’épithélium cilié si l’on est en présence de cellules ciliées.
Au niveau de la trachée, on décrit un épithélium pseudo-stratifié. En fait, cet épithélium est simple mais les cellules n’atteignent pas toutes le haut de l’épithélium. On observe des cellules différenciées (vieilles) et des cellules indifférenciées (jeunes) qui pourront remplacer les vieilles cellules.

C L’épithélium de la peau.
L’épiderme de la peau est un épithélium stratifié squameux, formé par des kératinocytes. Les cellules en contact avec la lame basale forment une assise génératrice pendant toute la vie de l’individu. Leur noyau est central, le cytoplasme bien colorable.
Les cellules, au dessous, sont reliées entre elles par des desmosomes, avec des filaments de cytokératine. Quand elles perdent le contact avec la lame basale, elles perdent l’aptitude à se multiplier et vont se différencier. Elles changent de forme et donneront les cellules de surface, mortes, aplaties, qui se détachent.
Les protéines synthétisées pendant la remontée interdisent les échanges et augmentent la protection. Quand il y a une destruction locale, ce sont les cellules de l’assise basale qui permettent la réparation des tissus.
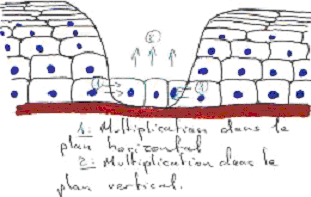
C’est par la lame basale qu’arrivent les facteurs de croissance intervenant dans la multiplication cellulaire.
Pour obtenir un tissu équivalent à l’épiderme, il faut cultiver des kératinocytes mais aussi des fibroblastes. On pourra alors avoir un tissu épithélial stratifié.
1 Les différenciations de l’épiderme
Les différenciations de l’épiderme vont pénétrer jusque dans le derme. Les poils ou les cheveux sont profondément implantés dans celui-ci ; ils ont une origine épidermique. Autour, on va trouver des cellules épithéliales formant un tissu stratifié. A la base, on a une papille de tissu conjonctif avec des filets nerveux.
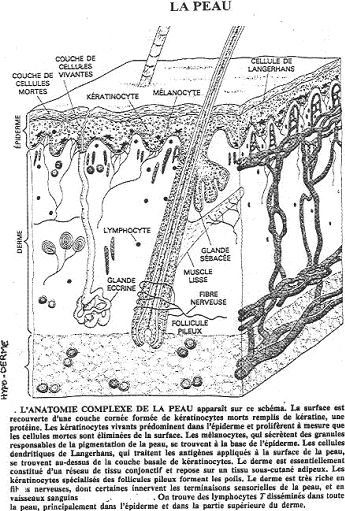
Une glande exocrine, la glande sébacée libère des lipoprotéines.
Un muscle lisse s’insère sur la formation pileuse et la rend mobile (muscle horripilateur).
Des glandes, formées d’un épithélium simple, pénètrent dans le derme : les glandes sudoripares.
Ces structures doivent leur maintien au tissu conjonctif sous-épithélial.
2 Les tissus conjonctifs.
La peau est un organe formé de deux tissus différents. Le derme comprend un tissu conjonctif lâche et un tissu formé de cellules séparées les unes des autres au sein d’une matrice extra-cellulaire.
Remarque : il existe d’autres tissus conjonctifs spécialisés dans la fonction de soutien : cartilage, os. Le sang est formé de cellules dans une matrice extra-cellulaire fluide.
3 Le derme.
a Cellules mises en jeu.
Le derme est formé de cellules qui sont à l’origine de la matrice extra-cellulaire (fibroblastes, adipocytes, …).
- Les adipocytes forment des plages dans la partie profonde du derme (l’hypoderme). Par leur fonction de stockage, ils interviennent dans l’homéothermie.
- Cellules émigrées du sang : lymphocytes B et T et plasmocytes. Les lymphocytes sont aisément reconnaissables par leur volume nucléaire important (noyau sphérique très colorable). Les plasmocytes sont des cellules actives dans la production d’Ig. Ces cellules présentent d’importantes modifications du cytoplasme. Elles ont un noyau sphérique, le cytoplasme est beaucoup plus abondant et contient des profils de REG pour la synthèse de protéines exportables, le golgi est bien développé. C’est dans le golgi que les protéines venant du REG sont modifiées (par glycosylation). Il part ensuite du golgi, des vésicules d’exocytose qui libèrent les Ig synthétisées à cause de la présence d’Ag.
- Les histiocytes. Ce sont des macrophages résidant. Ils ont de nombreux lysosomes dans lesquels est assurée la dégradation de particules étrangères.
- Les mastocytes. Ils contiennent dans leur cytoplasme de volumineux grains de sécrétion. Dans les grains sont accumulés des molécules de protéoglycanes (comme l’histamine). Leur exocytose est déclenchée par la présence (dans la matrice extra-cellulaire) de cellules étrangères à l’organisme. L’histamine attire les globules blancs : c’est une réponse du tissu à un traumatisme. Les mastocytes sont visibles en microscopie photonique si la coloration est adéquate.
- Les fibroblastes. Ils produisent la majorité des produits de la matrice. Ils sont souvent de petite taille, allongés. Autour de ces cellules, on voit de grandes fibres dans la matrice. Ce sont des cellules spécialisées dans la synthèse de fibres de collagène et de protéoglycanes.
b Les transformations des produits de synthèse des fibroblastes.
Les molécules de collagène : plusieurs sortes de gènes permettent la synthèse de nombreuses chaînes peptidiques qui sont les éléments de base des molécules de collagène mature.
Une molécule de collagène est une chaîne peptidique de grande taille (300nm de long). Cette chaîne a une séquence particulière : un acide aminé sur trois est une molécule de glycocol. Les autres acides aminés sont essentiellement des lysines et des prolines. On n’y trouve pas de tryptophane. Chaque molécule correspond à l’assemblage de trois chaînes peptidiques. On a une région en cordage dans la partie centrale ; les extrémités (N ou C) restent libres les unes les autres.

Les transformations. La glycosylation a lieu sur les molécules d’asparagine (avec une greffe de chaîne osidique) et dans les parties terminales, aussi bien en N qu’en C. à Les glycosylations permettent le maintien de l’écartement des chaînes. L’hydroxylation va porter sur les acides aminés. La lysine est transformée en hydroxylysine et la proline en hydroxyproline. Il y a donc des modifications importantes de la structure des chaînes de collagène. Les maladies de la peau sont généralement dues à des modifications des enzymes qui participent à ces réactions).
Ces molécules, une première fois transformées, vont dans le golgi où elles seront encore modifiées. Il va y avoir une évolution des motifs N-liés et des glycosylations nouvelles sur les groupements hydroxyles (à chaînes O-liées). Ces dernières modifications sont importantes pour la stabilité ultérieure.
Ces molécules peuvent alors être dirigées vers la membrane plasmique. Leur sécrétion se fait de manière continue, sans stockage intra-cellulaire : c’est une sécrétion constitutive. Au moment de l’exocytose, il y a une dernière modification moléculaire : il y a un clivage des extrémités N et C terminales è la molécule mature est uniquement composée de la région centrale du précurseur.
A l’extérieur, ces molécules s’assemblent et forment de grandes fibres visibles à la périphérie des cellules. L’assemblage se fait par formation d’un filament de fibrilles élémentaires.

Les striations parallèles sont claires et régulières.
Des liens intermoléculaires covalents renforcent ces assemblages et les fibres ainsi formées ont une solidité exceptionnelle à la traction comme les tendons (pour l’insertion des muscles). Ces tendons sont formés presque exclusivement de collagène. La résistance est due à la composition chimique et à l’assemblage.
Les molécules de collagène forment un réseau tridimensionnel à l’extérieur des cellules. Ce réseau permet à d’autres molécules (comme les protéoglycanes) de se stabiliser.
Les cellules libres peuvent même se frayer un passage à travers le réseau.
Le collagène formant ces fibres est appelé « collagène de type I ». Il est caractéristique des tissus conjonctifs lâches.
D’autres collagènes existent dans les tissus de soutien : la matrice extra-cellulaire est spécialisée à l’interface entre un tissu épithélial et un tissu conjonctif.
La lame basale, les tissus épithéliaux, le tissu conjonctif, les capillaires sanguins contiennent du collagène de type IV. Il a la particularité de ne pas subir de clivage des extrémités N et C au moment de l’exportation. De plus, il s’organise de manière à former des réseaux alvéolaires.
Ces réseaux se superposent les uns les autres et ils donnent une surface où les cavités ne sont pas obstruées : l’ensemble est un support pour les cellules qui y sont au contact.
Les cellules épithéliales établissent des interactions médiées par les intégrines et les fibres de fibronectine placées au niveau du collagène de type IV :
- il y a ancrage des cellules épithéliales.
- Il y a filtration des toutes les molécules dont la taille est inférieure à celle du réseau.
Dans cette lame, on trouve aussi des facteurs de croissance qui permettent la prolifération des cellules épithéliales. Il y a aussi des facteurs de différenciation cellulaire. Par exemple, l’épithélium intestinal est formé de cellules peu différenciées qui, au moment de leur formation et de leur migration, se différencient grâce aux facteurs rencontrés.
Les protéoglycanes sont aussi synthétisés par les fibroblastes. Ce sont des molécules formées d’un composant protéique sur lequel sont greffés (sur les résidus sérine) des chaînes en nombre et taille variable. Une chaîne est formée de glucose amino-glycane (ou GAG). Chaque chaîne a une longueur variable mais souvent élevée.
Les disaccharides répétés de ces chaînes sont composés d’un sucre aminé (glucose aminé) et d’un sucre acide. Ces disaccharides peuvent être répétés de nombreuses fois (10 à 150 résidus). Ces chaînes sont libres ou greffées sur des protéines. Quand elles sont greffées sur des protéines, on a des protéoglycanes.
Les GAG sont chargés (hydrophiles) et permettent de retenir l’eau et les substances dissoutes au sein des matrices qu’elles structurent.
Il existe des formes protéiques ou glycoprotéiques à distribution variable :
L’élastine : elle est dans la matrice extra-cellulaire sous-épithéliale des vaisseaux sanguins. Comme elle est hydrophobe, elle se replie sur elle-même. L’élastine va établir, avec les molécules du même type, des liaisons covalentes. L’assemblage se comporte, lors de l’étirement, comme un élastique. Quand l’élastine est en quantité importante, on a une bonne élasticité des parois.
La fibronectine : elle est formée de glycoprotéines de grande taille. Au niveau des lames basales, elle permet l’adhérence des cellules épithéliales. Elle se trouve aussi dans la matrice extra-cellulaire où elle joue un rôle important dans l’embryogenèse : les cellules en migration circulent selon les réseaux de fibronectine.
Les capillaires sanguins sont particulièrement abondants près de l’épiderme. Ils peuvent, en fonction des variations de leur calibre, participer à l’homéothermie. La vasoconstriction (pâleur de la peau) permet la diminution des pertes caloriques.
Les filets nerveux peuvent arriver jusque dans la région proche de l’épiderme où se différencient des structures sensorielles (bourgeons du tact).
La peau est un organe composé de deux tissus (derme et épiderme) qui sont irrigués, innervés et qui participent à la sensibilité et à l’homéothermie.
II Les Tissus Conjonctifs de soutien.
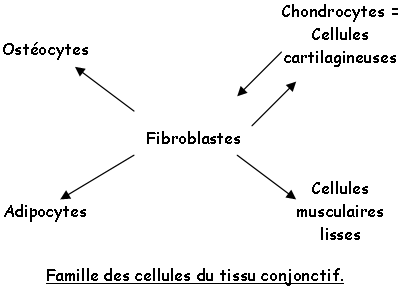
L’inter-conversion et la différenciation de ces cellules est réalisée par les hormones, les facteurs de croissance…
A Le cartilage.
Les chondrocytes sont des cellules productrices de collagène et de protéoglycane. Elles sont relativement sphériques. Leur taille est proche de celle d’une bactérie. Leur cytoplasme est riche en réticulum endoplasmique rugueux (pour la synthèse de protéines exportables) et le golgi est bien différencié (sert à la maturation des protéines).
Autour de ces cellules, on trouve une matrice extra-cellulaire avec de nombreuses fibrilles de petit calibre et courtes de longueur (collagène de type II).
Dans le réseau formé, se trouvent des protéoglycanes abondants et de grande taille. Ils sont caractéristiques des matrices extra-cellulaires de chondrocytes.
1 Constitution du cartilage.
Les gros assemblages de cartilage sont formés par la liaison sur l’acide hyaluronique (GAG), sur les placements de nombreuses molécules de protéoglycane (avec partie protéique et chaîne de glucose amino-glycane).
2 Nutrition du cartilage.
La structure du cartilage est importante dans sa physiologie car c’est le seul tissu vivant qui est sans capillaire sanguin. Il est pourtant constitué de cellules vivantes. Les cellules reçoivent les nutriments depuis les capillaires situés à l’extérieur du cartilage, via, les super-assemblages de protéoglycane.
Lors de la compaction du cartilage, l’eau et les substances dissoutes des assemblages, sont chassées vers le liquide articulaire et vers les vaisseaux sanguins. A la décompression, des fluides riches en nutriments pénètrent et sont mis à disposition des cellules pour leur nutrition. La vie de ce tissu dépend de ces mouvements mécaniques.
3 Constitution du cartilage.
Les cellules cartilagineuses se divisent et selon la disposition de ces divisions, on obtiendra deux types de tissus.
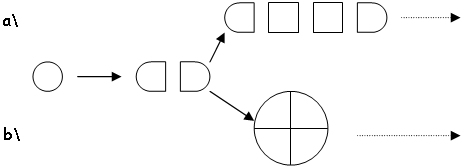
En a, les divisions amènent à des piles de cellules. C’est un groupe isogénique axial. On trouve ces divisions dans les zones d’élongation des os.
En b, on voit le cas des divisions dans la tête des os et dans la périphérie des vertèbres. L’origine est isogénique coronaire.
Remarque : c’est à partir de cartilage isogénique axial que se fait l’ossification endochondrale.
B L’os.
L’os est formé de cellules disposées dans une matrice extra-cellulaire : ce sont les ostéocytes. La matrice est appelée « osséine » ; elle est faite de collagène de type I. Elle contient des protéoglycanes et surtout, sert de support aux dépôts minéraux qui donnent la solidité de cette matrice.
Les ostéocytes ont des caractéristiques très proches de celles de chondrocytes : beaucoup de réticulum endoplasmique rugueux, un golgi bien visible è ce sont des cellules sécrétrices de protéines. A la périphérie, on trouve des fibres de grosse taille (collagène de type I). Quand on s’éloigne, la matrice devient minéralisée. Les cellules bénéficient d’un apport de nutriments en émettant des prolongements cytoplasmiques qui se dirigent vers les capillaires sanguins.
Autour des capillaires on voit une couronne de cellules qui envoient des prolongements. Les cellules lointaines envoient des prolongements sur les cellules précédentes (avec des jonctions ouvertes). è On a une organisation en rangées concentriques de cellules.
La coupe d’un os décalcifié montre une organisation avec plusieurs rangées de cellules (en cercle) qui est caractéristique des os longs : les systèmes de Havers.
L’os est un tissu vivant capable de se remodeler, de se refaire : c’est un tissu dynamique. On peut voir en coupe des systèmes fonctionnels et d’anciens systèmes non fonctionnels. Ce remodelage est possible grâce à l’intervention dans les os, d’une catégorie cellulaire particulière : les ostéoclastes. Ces cellules sont des cellules géantes, plurinucléées et qui présentent une polarité fonctionnelle. Ces cellules ont, à l’un des pôles, une grande surface de membrane formée de microvillosités. La face opposée est lisse. Malgré cette constitution, c’est une cellule libre.
Au pôle avant, il y a sécrétion d’hydrolases qui dégradent les protéines de la matrice osseuse, ce qui crée un passage pour l’entrée d’un capillaire. à On va avoir un nouveau système qui se développe à partir d’un système déjà formé. Cette capacité permet l’augmentation du diamètre de l’os et la soudure des fractures.
L’ossification endochondrale :
L’ébauche cartilagineuse n’est pas vascularisée. Il suffit qu’un capillaire sanguin pénètre, pour que se crée une zone d’ossification. Les ostéoclastes broutent le cartilage comme l’os. On va obtenir une région ossifiée de plus en plus grande, jusqu’à atteindre le plein diamètre.
La croissance en hauteur est possible grâce aux ostéoclastes qui dégradent une colonne de chondrocytes pour permettre l’entrée d’un nouveau capillaire.
III Le tissu musculaire, rapide rappel.

L’Appareil Digestif Mammifère.
La bouche joue un rôle dans la gustation, la mastication, l’imprégnation par la salive et la déglutition.
L’estomac intervient dans le brassage, l’acidification et dans l’attaque enzymatique.
L’intestin grêle collabore dans la digestion enzymatique, l’absorption des nutriments et la défense immunitaire.
I La bouche ou cavité buccale.
A La gustation.
La fonction de gustation est assurée par la langue.
La cavité buccale, chez les mammifères est revêtue d’un épithélium stratifié squameux. Au niveau du plancher buccal, une différenciation de ce plancher forme la langue. Cette langue est revêtue, sur la face supérieure, de différenciations particulières : les papilles. Ces papilles peuvent avoir des formes différentes. Les papilles filiformes sont les plus nombreuses et les plus petites. Elles ont un relief à symétrie axiale. Le relief des papilles facilite le transport des aliments vers les mandibules. Le tissu épithélial est protégé par un tissu conjonctif sous-épithélial qui forme une saillie.
Les papilles fungiformes sont nombreuses, au milieu des papilles filiformes. Elles ont une surface, au niveau de la cavité de la langue, relativement importante. Dans leur axe pénètre du tissu conjonctif sous-épithélial.
Les papilles circumvallées, très grosses, sont entourées par un sillon profond. Elles existent dans la partie postérieure de la langue et forment, chez l’Homme, le « V » lingual. Elles ont une grande surface de contact avec les aliments. Elles possèdent, sur leurs faces latérales, de nombreuses différenciations (bourgeons du goût) qui permettent la gustation par la présence de cellules sensorielles à leur niveau.
Les bourgeons du goût sont des différenciations sur la hauteur de l’épithélium. Le bourgeon du goût est en contact avec l’extérieur par une fossette : le pore gustatif. A l’opposé du pore, les cellules sont en contact avec la lame basale : ce sont de grandes cellules disposées comme les quartiers d’une orange. Ces bourgeons présentent deux catégories cellulaires :
- Les cellules sensorielles. Elles transforment les signaux chimiques perçus à l’apex, en signaux électriques transmis aux nerfs sensitifs. Elles sont maintenues par l’autre catégorie cellulaire.
- Les cellules de soutien. Elles maintiennent les cellules sensorielles.
Les aliments se déposent sur le pore gustatif et y restent le temps de la perception du goût. On trouve aussi ces bourgeons sur les papilles fungiformes mais moins fréquemment.
B Les glandes linguales.
A la base des papilles circumvallées, il y a arrivée du canal des glandes linguales, situées dans la partie profonde de la langue. Elles sécrètent de la salive, déversée au niveau du sillon périphérique de la papille : cette salive permet le nettoyage du sillon (retrait des aliments).
Ces trois types de papilles caractérisent la face supérieure de la langue. La face inférieure est revêtue du même type d’épithélium stratifié que la face supérieure mais sans papille. Les papilles ont pour fonction l’adhérence, l’humidification et la perception du goût.
Le transport des aliments est favorisé par la présence, dans le tissu conjonctif, de fibres musculaires striées, disposées selon tous les plans de l’espace (sections transversales, longitudinales…). Ces fibres permettent le déplacement volontaire de la langue. Elles vont traverser un tissu conjonctif lâche, comparable à celui du derme. Ce tissu conjonctif est bien vascularisé (par des capillaires sanguins). On peut observer des trajets nerveux à côté des papilles.
Dans la partie profonde de la langue, on remarque la présence, dans le tissu conjonctif, de glandes linguales. Elles peuvent être de deux types : les glandes muqueuses et les glandes séreuses. Elles sont faciles à distinguer par l’aspect des cellules.
- Les glandes muqueuses. Les cellules (les plus grandes) sont regroupées en acini et exportent les produits de sécrétion vers un canal de sécrétion, au centre (de la glande). Le noyau cellulaire est souvent plaqué contre la membrane basale et tout le cytosol est rempli de sécrétions (peu colorées en microscopie photonique).
Ces cellules produisent surtout des glycoprotéines et des protéoglycanes.
- Les glandes séreuses. Dans la même région de la langue, on trouve les glandes avec des cellules composant des acini. Leur noyau est sphérique, bien visible, dont le cytoplasme est plus colorable que les cellules des glandes muqueuses.
En microscopique électronique, on observe des grains de sécrétions contrastés, souvent hétérogènes. Les cellules produisent des sécrétions protéiques et parfois des glycoprotéines.
Les deux types cellulaires donnent une salive contenant des protéoglycanes, des glycoprotéines et des protéines (dont des hydrolases comme l’amylase). L’amidon est dégradé en chaînes d’amidon moins longues par l’amylase.
Les glandes salivaires débouchent dans le plancher lingual, sous la langue. Elles permettent la déglutition et l’entrée dans l’œsophage.
II L’œsophage.
L’œsophage est un conduit tubulaire.
On reconnaît dans la paroi, quatre tuniques caractéristiques de tout le tube digestif. Il y aura des variations des tuniques en fonction du niveau considéré. De l’intérieur vers l’extérieur, on distingue :
- La muqueuse.
- La sous-muqueuse.
- La musculeuse.
- La séreuse.
A La muqueuse œsophagienne.
La muqueuse est formée par un tissu épithélial stratifié squameux (comme dans la cavité buccale). Il repose sur une lame basale, elle-même est l’interface avec un tissu conjonctif lâche (ou chorion). L’ensemble (tissu conjonctif, lame basale, épithélium stratifié) est la muqueuse.
B La sous-muqueuse œsophagienne.
La séparation de cette muqueuse est souvent difficile à voir. On y trouve un tissu musculaire lisse : la musculaire de la muqueuse.
Dans le tissu conjonctif, on trouve parfois des glandes œsophagiennes qui sécrètent un fluide riche en protéoglycanes qui permettent, après exportation, l’humidification de la surface muqueuse ce qui facilite l’acheminement du bol alimentaire.
La sous-muqueuse est toujours formée de tissu conjonctif lâche, bien vascularisé, composant une assise relativement épaisse dans l’œsophage.
C La musculeuse œsophagienne.
Cette musculeuse est formée, dans son tiers supérieur, de muscles striés. Ailleurs (dans les deux tiers inférieurs), on n’aura que du muscle lisse. Les fibres sont disposées selon deux couches et sont plus ou moins obliques. On va distinguer une assise interne constituée de fibres circulaires et une assise externe formée de fibres longitudinales. Ce sont ces deux couches qui forment la musculeuse.
L’action conjuguée des deux couches musculaires permet la progression du bol alimentaire.
D La séreuse œsophagienne.
Cette séreuse est une tunique mince de tissu conjonctif.
L’œsophage joue un rôle majoritairement d’acheminement des aliments vers l’estomac.
III L’estomac.
L’estomac est une poche composée de trois parties : le cardia, le fundus et le pylore.
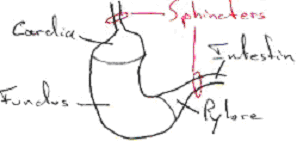
Le fundus a une fonction d’acidification et d’attaque chimique. Les aliments sont retenus dans l’estomac par des sphincters.
A La muqueuse.
L’épithélium de l’estomac est simple. La transition entre œsophage et estomac est abrupte. L’épithélium gastrique s’invagine (au niveau du fundus, principalement) pour former des glandes gastriques, serrées les unes contres les autres.
L’épithélium de surface (différent de celui des glandes) est constitué de cellules palissadiques, toutes égales entre elles. Elles possèdent, dans leur partie apicale, des grains de sécrétion riches en glycoprotéines colorées en rouge pourpre par le PAS (le PAS colore les glycoprotéines et les glucides).
Traitement au PAS :
On commence par traiter les coupes par un acide période puis par le réactif de Schiff. L’acide agit sur les cycles osidiques, en position 2 et 3 : il y a une oxydation qui fait apparaître les groupements OH. Le réactif de Schiff est formé de fuschine acide qui, lorsqu’elle rencontre du SO2, devient jaune pâle.
Le réactif se fixe sur les fonctions aldéhydes créées par oxydation.
Le rouge montre les glycoprotéines et les glucides (polysaccharides).
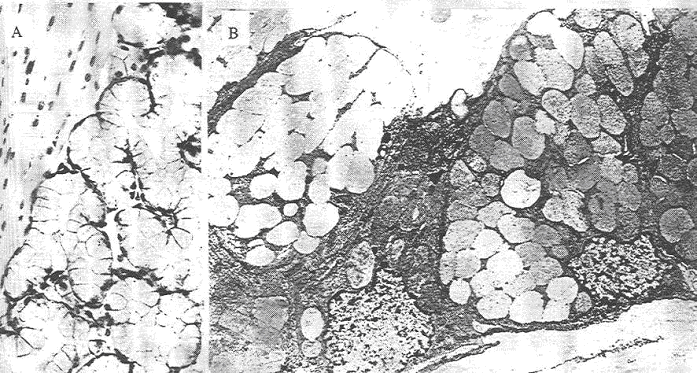
Les glandes gastriques contiennent plusieurs catégories cellulaires dont l’aspect et la fonction sont bien différents. On a trois types cellulaires :
- Les cellules principales sont plus importantes dans le fond des glandes.
- Les cellules bordantes (ou pariétales) sont plutôt dans la partie supérieure des glandes.
- Les cellules endocrines, les moins nombreuses, sont dispersées entre les deux autres types cellulaires.
Toutes les cellules des glandes participent à l’élaboration du suc gastrique.
1 Les cellules principales.
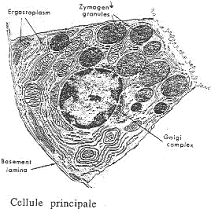
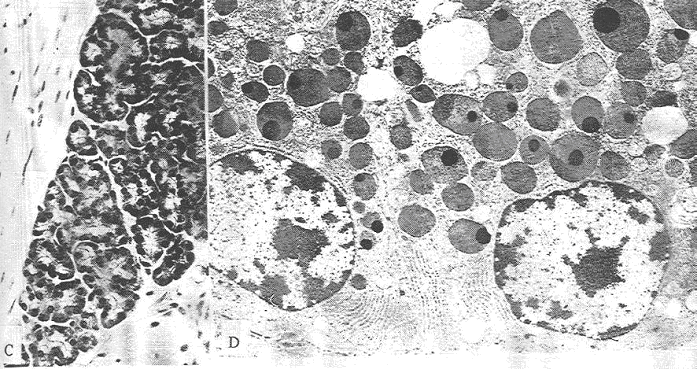
Les cellules principales produisent le pepsinogène, précurseur de la pepsine (protéine). Celle-ci est synthétisée dans le réticulum endoplasmique rugueux, bien développé dans la base de la cellule. Le golgi est aussi bien développé et il est à la base des grains de sécrétions, volumineux, qui s’accumulent près du pôle apical. Dans ces grains sont accumulés les précurseurs de la pepsine.
Ces cellules vont exocyter leurs grains de sécrétion de façon contrôlée. C’est l’entrée des aliments qui déclenche l’exocytose massive des grains de pepsinogène.
A pH < 7, le pepsinogène est transformé en pepsine (ensemble d’hydrolases capables d’agir sur les chaînes peptidiques). La pepsine est uniquement active à pH acide (le suc gastrique a un pH compris entre 1 et 3). Cette acidité vient de l’activité des cellules bordantes.
2 Les cellules bordantes ou cellules pariétales.
Les cellules pariétales libèrent l’HCl. Elles sont de forme généralement triangulaire. Leur aspect dépend particulièrement de leur activité.
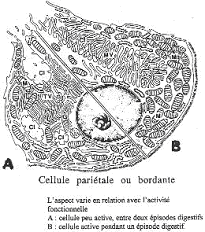
Au repos, ces cellules présentent un noyau sphérique avec de nombreuses mitochondries, peu de réticulum endoplasmique rugueux, un golgi développé mais de nombreux tubules et vésicules forment un réseau près de la membrane plasmique apicale. Cette partie de la membrane s’invagine de part et d’autre du noyau cellulaire pour former un canalicule intra-cellulaire. Les microvillosités sont peu nombreuses.
Quand ces cellules sont activées, elles changent de morphologie : il y a une augmentation considérable de membrane plasmique, due au développement de très nombreuses microvillosités. Là, on y trouve peu de réseau tubulo-vésiculaire.
Il y a une correspondance entre la quantité de réseau tubulo-vésiculaire et la taille de la membrane plasmique. En fait, ce réseau est de la membrane plasmique mise en réserve dans les cellules pendant les phases de repos. Elle sert à former les microvillosités pendant les phases actives.
La stimulation de ces cellules permet la sécrétion active d’HCL grâce à la présence de l’anhydrase carbonique (une enzyme) qui favorise la formation de protons et d’ions carbonates. Les protons sont libérés dans la lumière de la glande alors que les ions carbonates sont dirigés vers le sang.
L’hydrolyse incomplète des protéines à pH acide sera terminée dans l’intestin grêle.
3 Les cellules endocrines.
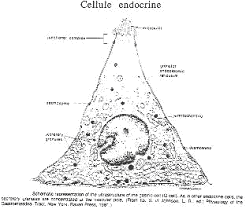
Ces cellules forment un ensemble producteur d’hormones capables de réguler l’activité des cellules exocrines. Ces cellules endocrines sont caractérisées par la présence de grains de sécrétion accumulés vers le pôle basal. Ces cellules sont souvent triangulaires, avec un faible contact apical et une surface développée au pôle basal, où se fait l’exocytose.
Il existe plusieurs catégories de cellules endocrines à effets variables :
- Les cellules à gastrine. Elles sécrètent des grains quand les aliments arrivent dans l’estomac. Le pH acide stimule ces cellules. La gastrine favorise la sécrétion de pepsine et d’HCl. Quand le pH est très bas, il y a un rétro-contrôle négatif sur la sécrétion de gastrine qui va s’arrêter.
B La sous-muqueuse et la musculeuse.
Sous l’épithélium, on trouve la muqueuse. Elle est constituée de tissu conjonctif lâche : le chorion. On y trouve aussi des capillaires sanguins qui circulent dans ce chorion, à proximité de l’épithélium (ils reçoivent les sécrétions hormonales).
La muqueuse se termine au niveau de la musculaire de la muqueuse, formée de muscles lisses.
La sous-muqueuse est composée de tissu conjonctif lâche.
La musculeuse est relativement épaisse et composée de deux couches de muscles lisses : circulaire interne et longitudinale externe.
C La paroi gastrique au niveau du cardia et du pylore.
De part et d’autre du fundus, la muqueuse présente quelques variations comme au niveau des glandes gastriques : on trouve moins de cellules pariétales et principales. On a surtout des cellules productrices d’ions (surtout au niveau du pylore).
Ces régions participent moins à la digestion.
Les sécrétions du pylore sont plus alcalines que celles du fundus et permettent de relever le pH du contenu stomacal avant l’entrée dans l’intestin grêle.
L’organisation de l’estomac est à peu près conservée chez tous les mammifères.
Chez les ruminants, le fonctionnement de l’estomac est différent : il existe plusieurs poches (la panse, le bonnet, le feuillet et la caillette). Panse, bonnet et feuillet correspondent (en histologie) à de l’œsophage de stockage et réalisent un brassage mécanique.
Avant l’entrée dans la caillette (la seule poche qui soit vraiment un estomac au sens histologique et fonctionnel) il y a des sécrétions gastriques à pH très bas qui contiennent des protéases caractéristiques de ce suc.
III Intestin grêle.
L’intestin grêle participe à la digestion des aliments qui arrivent de l’estomac. Il reçoit les sécrétions hépatiques et pancréatiques à fonctions digestives. Cet intestin possède des fonctions motrices et excrétrices (exocrines) par ses glandes muqueuses. Dans cette partie, il existe aussi des glandes sous-muqueuses.
La fonction majeure du grêle est l’absorption des nutriments. Des fonctions secondaires (mais importantes) sont par exemple la fonction excrétrice ou bien la fonction de défense immunitaire assurée par les plaques de Peyer.
A La fonction d’absorption.
La fonction d’absorption sera d’autant plus efficace que la surface d’absorption sera importante. C’est grâce à une grande surface épithéliale composée d’un épithélium simple qu’est assurée cette fonction.
Il existe des valvules qui sont des reliefs de grande amplitude qui soulèvent l’ensemble de la muqueuse. Il existe des reliefs de plus faible amplitude mais très nombreux : ce sont les villosités qui accroissent encore plus la surface de contact. Ce sont des évaginations à symétrie axiale présentes au niveau de la muqueuse. Ces évaginations sont recouvertes par un épithélium absorbant.
Le dernier dispositif permettant d’augmenter la surface de contact est formé des microvillosités. Ce sont des reliefs à symétrie axiale portés par des cellules épithéliales. Chaque cellule absorbante (entérocytes) possède de nombreuses microvillosités dont la hauteur est voisine de 1µm et de 0,1µm de diamètre.
Tous ces dispositifs vont permettre la mise en place d’une surface membranaire absorbante très importante. Chez l’Homme, le grêle mesure environ deux mètres de long et quatre centimètres de diamètres alors que la surface de contact est de 200m².
Les valvules permettent de multiplier par 3 la surface de contact, les villosités par 10 et les microvillosités par 20.
Le développement de ces structures est présent chez tous les mammifères.
B La paroi de l’intestin grêle.
Il apparaît pour la première fois au niveau du grêle, des microvillosités qui font parti de la muqueuse. L’épithélium s’invagine pour former les cryptes de Lieberkühn. Ces cryptes peuvent être simples ou ramifiées. Elles plongent dans le tissu conjonctif de la muqueuse. Ce tissu conjonctif muqueux va occuper l’axe des villosités et s’étendre jusqu’à la musculaire de la muqueuse (composée de muscles lisses).
Dans le tissu conjonctif sous-épithélial, on peut repérer, dans l’axe des villosités, un conduit chylifère ou lactéal. Dans ce tissu, sous l’épithélium, on trouve des capillaires sanguins.
La seconde tunique est la « sous-muqueuse » ; elle est formée de tissu conjonctif lâche parcouru de vaisseaux sanguins et de filets nerveux. Ces plexus nerveux ont leurs corps cellulaires à l’interface de la sous-muqueuse et de la musculaire (notamment les plexus de Meissner). Les corps cellulaires sont volumineux et partent en direction des autres cellules qui sont au même niveau.
Il vient ensuite la musculeuse, formée de deux tuniques musculaires lisses : une tunique musculaire circulaire interne et une tunique musculaire longitudinale externe. Entre ces deux tuniques existent les plexus d’Auerbach qui sont faciles à repérer entre les deux couches musculaires. Ils apportent les stimuli nécessaires à la contraction des fibres musculaires lisses.
C L’épithélium de la muqueuse.
L’épithélium de la muqueuse de l’intestin grêle est simple et palissadique. Il est composé majoritairement par des cellules absorbantes (entérocytes). Les autres cellules sont des cellules sécrétrices exocrines : cellules caliciformes à mucus ; elles sont moins nombreuses et disséminées dans les villosités.
Il existe aussi des cellules sécrétrices exocrines au niveau des cryptes de Lieberkühn : ce sont les « cellules de Paneth ». D’autres cellules à sécrétions endocrines constituent une population hétérogène dont le fonctionnement va réguler l’activité des autres catégories cellulaires mais également réguler les sécrétions de sucs et de sécrétions biliaires.
Toutes les cellules de l’épithélium de la muqueuse intestinale dérivent d’une cellule souche. Par multiplication, cette cellule donne une population de cellules indifférenciées, située dans la région des cryptes. Ces cellules ont la capacité de se multiplier en générant des cellules qui vont migrer soit vers le haut (villosité), soit vers le bas (crypte). Au fur et à mesure de leur migration le long de la lame basale, ces cellules vont se différencier :
- Si elles migrent vers le bas, on obtiendra des cellules de Paneth qui constituent le fond des cryptes.
- Si elles migrent vers le haut, elles donneront des cellules exocrines, ou caliciformes.
Il y a un renouvellement permanent de l’épithélium grâce à sa zone de multiplication.
1 Les entérocytes.
Les entérocytes sont des cellules hautes possédant des microvillosités particulièrement développées. Ces microvillosités forment une surface de membrane plasmique considérable. Elles possèdent des filaments d’actine qui se prolongent à l’intérieur du cytoplasme apical ; il n’y a pas d’organite dans cette région. Les microfilaments entrent en relation avec des microfilaments associés à la ceinture d’adhérence. Il apparaît à ce niveau un feutrage de surface avec des microfilaments : terminal web. On peut détecter la présence de myosine dans cette surface. Cette dernière, avec l’actine, rend mobiles les microvillosités. Ceci a l’avantage de permettre un renouvellement du fluide intestinal et faciliter son absorption.
En dessous, on trouve un réseau de réticulum endoplasmique lisse puis de réticulum endoplasmique rugueux. En dessous vient le chondrium (les mitochondries). Les mitochondries sont dans deux régions : près de l’apex et près de la base.
L’appareil de Golgi occupe une position supra-nucléaire. Le noyau occupe le tiers inférieur de la cellule. En région basale, on retrouve du réticulum endoplasmique rugueux et des mitochondries.
L’entérocyte repose sur une lame basale et est toujours placé à proximité d’un capillaire sanguin. La paroi du capillaire est très mince, afin de favoriser les échanges.
Les cellules sont moins étroitement associées à leur base et il est fréquent de voir s’infiltrer des lymphocytes entre les cellules.
2 Les cellules caliciformes à mucus.
Ces cellules sont nombreuses sur la face latérale des villosités. Elles sont réunies aux autres cellules par des jonctions fermées. Le noyau est basal avec à proximité, du réticulum endoplasmique rugueux très développé et un appareil de Golgi de grande taille. Ce type cellulaire synthétise des protéoglycanes et des gluco-amino-glycanes. Les grains vont s’accumuler. Ces régions sont fortement colorables au PAS.
Les grains de mucus vont, par exocytose, répandre le mucus à la surface de l’épithélium. Ses composants ont un rôle protecteur par rapport aux enzymes digestives de la lumière intestinale.
3 Les cellules de Paneth (exocrine).
Les cellules de Paneth occupent des diverticules. Elles sont caractérisées par leur forme globalement triangulaire. On note la présence de grains de sécrétion de nature protéique dans la région apicale. Ces cellules, à exocytose contrôlée, produisent des composés tel que le lysosome intestinal à propriétés bactéricides. La sécrétion permet de nettoyer la lumière des cryptes.
4 Les cellules endocrines : cf. TP.
5 L’intestin grêle est divisé en trois régions :
- Le duodénum : Les villosités y sont les plus trapues et y sont présentes les glandes de Bruner. Ces glandes, par leur acini exocrines, sécrètent un fluide à pH alcalin qui se déverse au niveau de la muqueuse du duodénum. Ce fluide permet de neutraliser le chyme alimentaire encore acide à la sortie de l’estomac.
- L’iléon : La musculeuse y est la plus épaisse.
- Le jéjunum.
D Fonctions digestives.
A la sortie de l’estomac, les protéines ont été attaquées par la pepsine et les glucides ont subit un début d’hydrolyse par l’amylase salivaire.
Pour obtenir l’hydrolyse complète des nutriments vont intervenir les sécrétions pancréatiques et hépatiques. C’est dans le duodénum que débouchent ces sécrétions (pancréatiques et biliaires).
Le pancréas sécrète de nombreux enzymes sous forme de précurseurs. Parmi ces enzymes, on trouve la trypsine (sous forme de trypsinogène) et la chymotrypsine (sous forme de chymotripsinogène). Ces composés agissent sur les aliments en les scindant en chaînes plus courtes : dipeptides ou tripeptides. En plus des protéases, on aura des lipases et des amylases.
Finalement, on a une trentaine d’enzymes qui s’attaquent aux protéines, aux glucides, aux lipides mais aussi à l’ADN et à l’ARN. Ces enzymes sont déversées dans le duodénum où leur action ne peut se faire qu’à pH neutre.
La lipase pancréatique agit sur les particules alimentaires telles que les triglycérides et son action n’est possible que grâce à la présence de sels biliaires qui vont permettre d’émulsifier les graisses en particules plus petites, attaquables par les lipases. Il y a alors obtention de monoglycérides et même d’acides gras.
Les amylases pancréatiques continuent l’attaque des polysaccharides. Cette attaque conduit à la formation de disaccharides (exemple : le maltose).
Les cellules absorbantes contiennent, au niveau de leur membrane, des protéines à fonction enzymatique. On trouve des protéines, sur les microvillosités, assurant la transformation des disaccharides en oses qui sont les molécules assimilables ; mais aussi des protéases (même localisation), et particulièrement des dipeptidases, qui convertissent les dipeptides en acides aminés assimilables. Le résultat conduit :
- A des oses pour les sucres ;
- A des acides aminés pour les protéines ;
- A des monoglycérides et des acides gras pour les lipides.
L’entérocyte est capable d’absorber les petites molécules. Pour l’absorption des oses, il existe des protéines synports sur la membrane des microvillosités : les oses sont absorbés en même temps que du sodium et cela, en raison de la polarité de l’épithélium. Il existe un flux de sodium à travers la cellule : les protéines synports font entrer le sodium alors que la pompe Na/K, sur la surface latéro-basale, va dépolariser l’entérocyte.
Les lipides, hydrophobes, n’ont pas le même comportement. Ils vont pénétrer par liposolubilité au niveau de la membrane apicale. On ne voit pas d’endocytose de lipides. Toutefois, dans la phase aqueuse du cytosol, ces lipides vont rencontrer le réticulum endoplasmique lisse où se trouvent les enzymes permettant l’estérification, qui, au final, permet la formation de triglycérides. Dans le réticulum endoplasmique lisse, on voit apparaître des gouttelettes lipidiques, sombres : ce sont les chylomicrons, obtenus par estérification.
Le réticulum endoplasmique lisse va acheminer ces chylomicrons vers le réticulum endoplasmique rugueux qui est le lieu de synthèse de protéines et particulièrement, de protéines lipophiles (produites par les entérocytes). Du réticulum endoplasmique rugueux, les chylomicrons transitent vers le Golgi et là, vont s’accumuler sous forme de grains de sécrétion. Ces derniers vont être exocytés dans l’espace intercellulaire ; ils sont ensuite acheminés vers la lame basale puis vers le tissu conjonctif sous-épithélial.
Cas particulier : Absorption chez les mammifères nouveaux-nés.
Les sécrétions pancréatiques et même les sécrétions de l’estomac ne sont pas encore efficaces pour l’hydrolyse du lait. Il arrive donc non dégradé au niveau de l’intestin grêle et est absorbé par un mécanisme d’endocytose. On peut voir de très nombreuses petites vésicules se former à partir de dépressions. Les vésicules plus grandes sont dirigées vers la partie supérieure du noyau.
Si expérimentalement, on introduit une protéine comme la péroxydase, on voit que la protéine rentre par endocytose, circule par un réseau tubulo-vésiculaire vers les lysosomes où elle sera dégradée, ce qui produit la libération d’acides aminés. Ces derniers seront dirigés vers sang. Les IgG du lait maternel pénètrent par endocytose, passent par le réseau tubulo-vésiculaire et finissent dans le sang. Ces Ig permettent un renforcement des capacités de défense immunitaire du nouveau-né.
IV Le gros intestin.
Le gros intestin présente des villosités intestinales. Sa muqueuse est composée d’un épithélium simple, palissadique qui s’invagine pour donner des glandes de Lieberkühn. Au niveau de ces glandes existent de nombreuses cellules caliciformes à mucus.
La muqueuse va jusqu’à la musculaire de la muqueuse.
La sous-muqueuse est particulièrement importante dans le gros intestin.
Les cellules à mucus favorisent le transit des résidus de l’absorption.
On a des cellules particulières : les cellules M. Ces cellules sont placées entre les entérocytes et leur forme dégage une cavité où viennent migrer des lymphocytes et des macrophages. Les cellules M sont capables de faire de l’endocytose et permettent le transit de molécules (antigéniques) jusqu’aux cavités existant dans les espaces intestinaux. Ces cellules M sont présentes dans l’iléon et dans tout le gros intestin (colon et rectum). On trouve fréquemment des plaques de Peyer qui sont des nodules contenant des lymphocytes participant aux fonctions de défense.
Le gros intestin a pour fonction essentielle d’héberger la flore intestinale. Ces microorganismes participent à la dégradation de composés non hydrolysés, comme par exemple la cellulose végétale. Par ailleurs, ce segment permet une forte réabsorption de l’eau, ce qui rend le contenu intestinal de plus en plus compact. A ce niveau sont excrétés les composés non hydrolysés.
La musculature favorise le transport jusqu’au rectum. Dans la partie postérieure, cette musculature est composée de muscles striés, ce qui permet une excrétion volontaire.
L’Appareil Digestif Non Mammifère.
Chez les cyprinidés, il n’y a pas d’estomac : le tube digestif est simple. Il est composé d’une cavité buccale, d’un pharynx et d’un intestin.
Les poissons comme la perche, le poisson-chat, ont un estomac.
I Cavité buccale et pharynx.
La cavité buccale et le pharynx sont en continuité. Dans la cavité buccale arrivent les aliments pour le tube digestif mais aussi l’eau pour les branchies. Le pharynx est la région constituée par les branchies.
Les aliments vont de la cavité buccale à l’œsophage alors que l’eau va de la même cavité buccale aux branchies puis ressort en arrière de l’opercule.
Les branchies ont un axe cartilagineux pourvu d’épines latérales qui s’interpénètrent entre branchies et forment un filtre qui retient les aliments, tout en laissant passer l’eau.
La région antérieure a un épithélium stratifié dont les cellules supérieures sécrètent un mucus. Elles peuvent parfois être ciliées.
Certains poissons ont des dents (ou odontoïdes) : ce sont des parties riches en cartilage dont l’origine embryonnaire est différente de celle des autres vertébrés. Ces « dents » sont réparties de manière variable, dans la cavité buccale ou pharyngienne, selon les espèces. Chez les carpes, il n’y a que des dents pharyngiennes, composées d’une pièce dorsale et de deux pièces ventrales, qui réduisent les aliments en fines particules.
L’œsophage est un conduit court dont la paroi est constituée d’un épithélium stratifié, comparable à celui de la cavité buccale (avec plusieurs catégories de cellules à mucus). Les cellules à mucus servent à faciliter le transit vers la suite du tube digestif.
II Les cyprinidés.
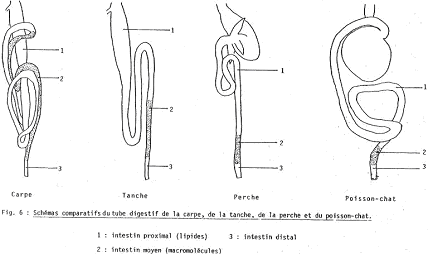
En aval de l’œsophage, on a un intestin dont la particularité est de voir son calibre diminuer de la partie proximale à la partie distale : il n’y a pas de différenciation morphologique pour la distinction de niveaux.
La paroi de l’intestin apparaît plus simple que chez les mammifères. On y trouve trois tuniques et non quatre. De l’extérieur vers l’intérieur, on trouve : la séreuse, la musculeuse (avec deux couches de muscles lisses à longitudinale externe et circulaire interne) et la muqueuse dont l’épithélium décrit des villosités qui ne sont que des replis ayant un plan de symétrie et non un axe. Elles permettent d’augmenter la surface de contact mais sont inférieures aux vraies villosités.
L’épithélium est simple, sans crypte de Lieberkhün. Il repose sur un tissu conjonctif lâche et chez certaines espèces, il existe des chylifères dans le tube digestif.
Il n’y a pas de musculaire de la muqueuse, donc, pas de sous-muqueuse. Dans ce tissu conjonctif, on voit fréquemment, à l’emplacement où se trouverait la musculaire de la muqueuse, des fibres de collagène qui forment le stratum compactum (faisceaux relativement denses).
L’épithélium palissadique est formé de trois catégories cellulaires :
- Les entérocytes (en majorité).
- Les cellules caliciformes à mucus, disposées aléatoirement.
- Les cellules endocrines servant à la régulation du fonctionnement des entérocytes.
- Il n’y a pas d’équivalent des cellules de Paneth.
L’organisation ultrastructurale des entérocytes permet de distinguer trois régions : l’intestin proximal, l’intestin moyen et l’intestin distal (court segment).
1 L’intestin proximal.
Les entérocytes ressemblent à ceux des mammifères. Le pôle apical comprend des microvillosités régulières. Il n’y a pas d’endocytose entre les microvillosités. Si les animaux sont alimentés, on voit que les entérocytes assurent l’absorption des lipides et possèdent un réticulum endoplasmique lisse.
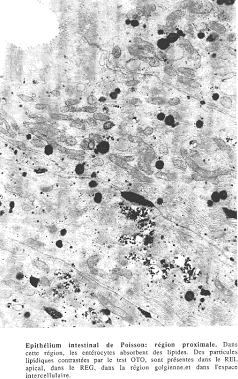
Ces lipides sont appelés « lipides particulaires » car la taille des globules est toujours inférieure à celle des chylomicrons.
Ces lipides passent du réticulum endoplasmique lisse au réticulum endoplasmique rugueux puis dans le golgi d’où ils iront dans des grains de sécrétion, capables d’exocyter leur contenu dans l’espace intercellulaire. La localisation est équivalente à celle des chylomicrons. Les lipides passent ensuite dans les capillaires sanguins après avoir passé la lame basale.
Ces lipides ont subit l’action des sels biliaires et des sucs pancréatiques. C’est dans la partie supérieure de l’intestin que débouche le canal cholédoque. Les lipides, ainsi transformés sont de petites molécules capables de passer la membrane plasmique.
Dans ces cellules, il existe aussi des lipides étalés qui apparaissent après les repas, par estérification. Les lipides ne se trouvent que dans les entérocytes. Ils devraient représenter une forme de stockage cellulaire momentanée des lipides. Il est possible de les convertir en lipides particulaires ; ce qu’il se passe en période post-repas (jeûne).
2 L’intestin moyen.
L’épithélium change d’organisation abruptement et les entérocytes de l’intestin moyen sont capables de faire de l’endocytose de macromolécules. Un réseau de tubules et de vésicules existe et leur formation est de type lysosome. Ils sont en position supra-nucléaire. Les protéines pénètrent par endocytose, circulent par les tubules et vésicules vers les lysosomes pour y être dégradées. On assiste à une digestion intra-cellulaire des protéines qui compense le manque d’une hydrolyse intra-luminale.
3 L’intestin distal.
Les entérocytes de cet intestin sont différents des autres. Le plateau des microvillosités est moins bien structuré (microvillosités de petite taille, dispersées) que chez les précédents. La membrane plasmique s’invagine dans la région péri-nucléaire, sur les faces latérales et basales. Ces invaginations sont souvent accolées à des mitochondries, nombreuses dans la région basale, entre les replis membranaires.
Il y a des échanges transmembranaires très actifs.
Ces cellules peuvent travailler à des transports d’eau ou d’ions. Elles sont comparables aux cellules des tubes collecteurs du rein : rôle dans l’osmorégulation.
III Les poissons à estomac.
Chez ces poissons, l’estomac est une poche volumineuse en deux parties. A la suite de l’œsophage, on a l’estomac antérieur, l’estomac postérieur puis l’intestin.
D’un point de vue histologique, l’estomac est composé par trois tuniques (de mêmes noms que précédemment). La différence entre estomac antérieur et postérieur réside dans le fait que l’estomac antérieur possède des glandes gastriques, à la différence du postérieur.
Il existe un stratum compactum et la musculeuse (circulaire interne, longitudinale externe). Cette tunique est beaucoup plus épaisse dans l’estomac postérieur.
L’épithélium contient des cellules de surface hautes et étroites. Elles sont toutes semblables entre elles et présentent des grains de sécrétion PAS positifs à leur apex (comparable à celle des mammifères).
Les glandes contiennent un type cellulaire : des cellules totipotentes qui sécrètent l’HCl et des protéases de type pepsine. Ces cellules ont à la fois des caractères de cellules à HCl (nombreuses mitochondries et réseau développé de tubules et de vésicule quand la cellule est au repos) et de cellules productrices de protéines (réticulum endoplasmique rugueux bien développé dans le pôle basal, golgi de grande taille qui donne des grains de sécrétion volumineux). Les grains contiennent des protéases type pepsine : ce sont des grains de zymogène.
L’estomac postérieur assure un brassage mécanique très efficace grâce à la forte musculature, les protéines ayant subi une hydrolyse à pH acide.
Dans l’intestin, l’hydrolyse se poursuit dans, au moins, deux régions.
- L’intestin antérieur : il porte de nombreux diverticules (des centaines) appelés « cæcums pyloriques » En coupe transversale, la paroi est comparable à celle des cryptes, comme dans l’intestin proximal de la carpe. La coupe transversale des cæcums montre un organe comparable à celle de l’intestin avec un calibre réduit. Cette partie assure l’absorption des lipides alimentaires. Les cæcums fonctionnent comme l’intestin proximal : ce sont juste des dispositifs permettant l’accroissement de la surface absorbante.
- L’intestin postérieur : il possède une même organisation que chez les cyprinidés ; les entérocytes peuvent endocyter les protéines pour assurer une digestion intra-cellulaire. La digestion par l’estomac n’est pas suffisante pour digérer les protéines en acides aminés. Les macromolécules doivent être endocytées.
Les valvules spirales sont des dispositifs intestinaux (dans le conduit) qui forment des replis à l’intérieur de la lumière. Ce sont des reliefs du stratum compactum qui soulèvent toute la muqueuse. Elles permettent une forte augmentation de la surface épithéliale.
IV Les amphibiens/reptiles.
Chez les amphibiens adultes existe sur le plancher buccal, une langue mobile et différenciée. Elle sert souvent à capturer les proies et se replie vers l’extérieur, ancrée par l’avant, pour exposer la face inférieure pendant la capture.
Bouche fermée, la face inférieure correspond à la face supérieure des espèces plus évoluées.
La langue est formée d’un épithélium stratifié, lisse, sur la face supérieure. Cet épithélium s’invagine sur la face inférieure pour former les papilles. L’épithélium est riche en cellules à mucus et contient parfois des cellules ciliées. Le mucus se trouve sur toute la surface de la langue et sert à agglutiner les proies.
Sous l’épithélium, on trouve un tissu conjonctif dans lequel sont disposées des fibres musculaires striées en faisceaux transversaux, longitudinaux… (dans toutes les directions de l’espace).
Il vient ensuite l’œsophage, court, puis un estomac, qui, lorsqu’il est différencié, a une organisation comparable à un estomac de poisson : la région antérieure possède des glandes gastriques ; la région postérieure est sans glande. A la différence des poissons, on trouve une musculaire de la muqueuse (donc, 4 tuniques).
Les glandes gastriques comprennent des cellules totipotentes pour la sécrétion d’HCl et de protéases (type pepsine).
L’intestin est un conduit à calibre qui se rétrécie lorsque l’on s’éloigne de l’estomac. La paroi, en coupe, montre quatre tuniques classiques.
La muqueuse est formée d’un épithélium simple, donnant des villosités à symétrie axiale. Entre les villosités, à leur base, existent des nids cellulaires (les massifs sous-épithéliaux) capables de se multiplier pendant la vie de l’animal. Ces cellules colonisent l’épithélium des villosités et permettent un renouvellement de cet épithélium (comme les cryptes de Lieberkühn).
Remarque : chez les poissons, il y a multiplication des cellules du fond des replis.
Comme peu de travaux ont été réalisés chez la grenouille, on n’en sait assez peu.
Chez les reptiles, on retrouve les mêmes caractéristiques que chez les batraciens.
V Les oiseaux.
On retrouve chez les oiseaux des caractères bien plus comparables à ceux des mammifères. Les différences sont dues à la nature des éléments ingérés. Les oiseaux sont des granivores (et consomment donc des graines).
La cavité buccale est revêtue par un épithélium stratifié squameux (comme les mammifères). Il existe une langue peu ou pas mobile sur le plancher buccal. Cette langue contient dans sa partie postérieure, des glandes salivaires. La salive va humidifier les graines.
L’œsophage est un conduit relativement long. Sur son trajet, l’œsophage se différencie en une poche, le jabot. Cette poche a la même structure mais est plus riche en glandes sécrétrices. L’épithélium, stratifié squameux, repose sur le tissu conjonctif sous-épithélial, avec des cellules sécrétrices formant les glandes oesophagiennes. Ces cellules produisent, chez certaines espèces, une sécrétion régurgitée par les parents dans le bec des nouveaux-nés, afin de les nourrir.
On trouve aussi une musculaire de la muqueuse qui sépare le tissu conjonctif muqueux du tissu conjonctif sous-muqueux. Vient ensuit la musculaire, classique, et une séreuse.
L’estomac est formé de deux parties :
La partie antérieure est appelée stomodeum ou ventricule succenturié. C’est la région pourvue de glandes qui produisent les sécrétions acides et les protéases (type pepsine).
La partie postérieure est le gésier. Il ne contient pas de glande gastrique. Il assure la digestion mécanique des graines déjà attaquées par les sucs dans la partie antérieure. La muqueuse différencie des glandes épithéliales sécrétant des protéines (la koïline) : des scléroprotéines qui forment une couche épaisse et protectrice.
Le tissu conjonctif occupe l’espace sous-épithélial. La musculaire est constituée de fibres lisses.
La sous-muqueuse est peu épaisse, formée de tissu conjonctif.
La musculeuse est très épaisse, formée de deux couches (circulaire interne, longitudinale externe). Elle assure ainsi un broyage mécanique de très grande efficacité.
L’intestin est chez les oiseaux, très comparable à l’intestin grêle des mammifères.
Les Glandes Annexes et le Foie.
I Chez les mammifères.
A Organisation générale.
Le foie reçoit le sang par la veine porte. Il est en relation avec la circulation générale avec l’artère hépatique (une branche de l’aorte). Le sang repart vers la veine cave par les veines sus-hépatiques. Le foie est un organe qui reçoit les aliments directement après leur assimilation par le tube digestif. Il les stocke et modifie les nutriments avant de distribuer ses produits de synthèse vers le sang (c’est une glande endocrine) ou par les canaux biliaires (glande exocrine). è Le foie est une glande mixte.
La majorité des cellules qui composent le foie sont des hépatocytes (ou cellules principales du foie). Ce sont de grosses cellules, en travées, associées les unes aux autres par des jonctions fermées. Les travées cellulaires sont organisées en cordons rayonnant en direction de la veine centro-lobulaire. Cette veine occupe le centre du lobule hépatique.
Les hépatocytes ne sont pas les uniques composants du foie. On trouve les cellules Kuppfer, fonctionnant comme des macrophages, fixées à la paroi des capillaires sanguins. On trouve aussi une faible proportion de cellules de Ito (des lipocytes). Ces lipocytes sont le lieu de mise en réserve de vitamines liposolubles (A et B). On trouve aussi les cellules endothéliales de la paroi des capillaires.
B La cellule hépatique.
A la périphérie du lobule, on peut voir des sections de vaisseaux sanguins : on a une branche de la veine porte et une de l’artère hépatique. On a aussi un canal biliaire collectant les sécrétions endocrines du lobule. Le sang rentre dans les sinusoïdes (les capillaires du foie) ayant la particularité d’avoir une paroi très mince. Les sinusoïdes vont se diriger vers la veine centro-lobulaire.
Chaque cellule hépatique expose au moins deux faces au capillaire sanguin (pôle basal des cellules). Les microvillosités se trouvent sur la face tournée vers les capillaires. Il y a souvent endocytose sur ces faces là : il y a de nombreux échanges entre les cellules et le sang. Les microvillosités vont vers l’endothélium des sinusoïdes mais sont séparées par l’espace de Disse. Dans cet espace, on voit souvent des particules comme des lipoprotéines de basse densité (le foie intervient dans la régulation des lipides).
Le pôle basal est séparé du pôle latéral par une jonction fermée réunissant les cellules entre elles. A l’opposé se trouve deux cellules, des canalicules biliaires (à diamètre de petite taille) formées par la membrane plasmique de deux cellules contiguës et correspondent au pôle apical des hépatocytes.
Au centre de la cellule, on a un noyau sphérique dont la taille peut varier chez chaque hépatocyte : la quantité d’ADN montre que si les cellules diploïdes sont majoritaires, des cellules sont tétraploïdes, voir même octoploïdes.
A la périphérie du noyau, on a un réticulum endoplasmique rugueux développé et plusieurs systèmes golgiens sont visibles près des pôles biliaires. Les mitochondries sont nombreuses et leur distribution est homogène (environ 1000 mitochondries par hépatocyte). Le réticulum endoplasmique lisse est bien développé près du pôle basal. Il y a souvent des particules de glycogène qui sont proches du réticulum endoplasmique lisse et à elles seules, elles sont un indice pour identifier un hépatocyte. Elles sont de grande taille et souvent opaques aux électrons : ce sont des particules a (glycogène couplé à d’autres oses). Ailleurs, les particules sont plus petites : ce sont les particules b.
Quand les cellules sont soumises à un jeune prolongé, les particules a se dégradent et donnent les particules g, très petites qui persistent et pourront servir lors d’une reprise alimentaire pour former de nouvelles particules a. Dans ces particules, il existe le glycogène mais aussi les enzymes nécessaires à sa synthèse et à sa dégradation.
Les gouttelettes lipidiques sont une autre forme de réserves d’énergie.
C L’épithélium.
L’endothélium est perforé, ce qui accroît les échanges.
Un acini hépatique est différent d’un lobule : c’est un ensemble fonctionnel se trouvant de part et d’autre de la région péri-lobulaire jusqu’au centre des veines centro-lobulaires appartenant à deux lobules différents.
Dans l’axe de l’acinus arrivent les vaisseaux amenant les nutriments et le sang oxygéné. Le sang draine les hépatocytes vers la veine centro-lobulaire et vient se charger de nutriments.
La proximité de l’arrivée des vaisseaux est un lieu de stockage ; on trouve des zones en auréole moins chargées en nutriments que la région centrale de l’acinus.
D Les hépatocytes.
1 Fonction exocrine.
Les hépatocytes sont le lieu de synthèse des sels biliaires, du cholestérol et des lécithines qui composent la sécrétion biliaire. Les sels sont formés au niveau du réticulum endoplasmique lisse par un dérivé de la cystéine (la taurine) et le glycocol conjugué. Le cholestérol est synthétisé au niveau du réticulum endoplasmique lisse dont la synthèse se fait à partir d’acides gras. Les pigments biliaires viennent d’un dérivé d’éléments comme l’hémoglobine, dégradée en bilirubine.
Ces sécrétions sont acheminées, pendant leur production, vers le pôle apical pour l’exocytose. Les canalicules biliaires circulent entre les hépatocytes, en direction des canaux biliaires périlobulaires. C’est au niveau de la vésicule biliaire que s’accumule la sécrétion, en l’attente d’un repas. En réponse à des stimuli hormonaux, la bile est déversée pour atteindre le duodénum. Elle participe à l’émulsion des triglycérides alimentaires.
Les cellules épithéliales de l’intestin recaptent les dérivés des sels biliaires qui seront recyclés vers la vésicule biliaire après un passage par les hépatocytes.
2 Fonction endocrine.
Il y a synthèse de différentes protéines contenues dans le plasma (fibrinogène, albumine et globuline) qui sont sécrétées de manière continue vers le pôle vasculaire des cellules.
Les fonctions endocrines participent au métabolisme intermédiaire.
Pour les glucides, il y a capture du glucose alimentaire après le repas, pour le stockage en glycogène (donc, diminution de la glycémie). Quand cette glycémie est trop basse, il y a libération de glucose à partir du glycogène. La régulation de la glycémie est sous contrôle des hormones pancréatiques endocrines.
Dans le cas du métabolisme lipidique, après un repas, les lipides alimentaires sont transportés sous forme de chylomicrons en direction des vaisseaux lymphatiques (les chylifères). Ces derniers entrent en relation avec le système vasculaire au niveau de la veine sous-clavière. Les lipides sont distribués aux cellules adipeuses pour le stockage. Les cellules endothéliales du système sanguin prélèvent, par des lipases membranaires, différents composés des chylomicrons. Les composés restant arrivent aux hépatocytes qui absorbent à leur tout des particules du sang circulant pour les modifier.
Dans le sang circulant, on peut trouver : des chylomicrons de grande taille, des particules de taille inférieure à celle des chylomicrons qui contiennent toutes des lipides et protéines (triglycérides, cholestérol, phospholipides) mais dont les proportions dans les différents composés varient. Le cholestérol HDL est de haute densité alors que le LDL et le VLDL sont de faible densité. On les trouve surtout dans le sang.
Le foie est le lieu de production du cholestérol. Il prélève dans le sang circulant des particules (comme l’HDL) à teneur faible en cholestérol, les modifie en ajoutant du cholestérol et des triglycérides après le repas. On a alors du VLDL, qui, sous forme circulante, est utile aux autres tissus qui, en le captant, utilisent le cholestérol et les triglycérides contenus.
Deux catégories cellulaires savent synthétiser du cholestérol : les hépatocytes et les cellules à hormones stéroïdes. Elles fournissent le cholestérol à toutes les autres cellules qui l’utilisent.
En dehors des phases de repas, les hépatocytes synthétisent du cholestérol à partir d’acides gras (et donnent du VLDL).
Le foie est l’élément régulateur de la glycémie et un lieu de synthèse du cholestérol. Il intervient aussi dans les fonctions de détoxication en transformant ces composés (toxiques) en composés non toxiques, éliminés avec les sécrétions biliaires.
E Cellules de Kuppfer et cellules de Ito.
Les cellules de Kuppfer sont aisément reconnaissables par leur position, leur forme générale allongée et leur contenu souvent riche en fer. Elles peuvent phagocyter les cellules mortes et dégradent l’hémoglobine, ce qui donne les éléments riches en fer des sels biliaires.
Dans certaines anémies, ces cellules sont chargées en fer plus qu’à la normale.
Les cellules de Ito sont des cellules chargées en lipides servant à stocker des vitamines liposolubles (A et D).
II Le foie chez les non mammifères.
A Chez les vertébrés inférieurs (poissons).
Le foie est toujours composé d’hépatocytes de grande taille mais il n’y a pas d’organisation en lobule. Ces cellules sont en travées anastomosées. De plus, à l’intérieur du foie, on peut trouver du pancréas exocrine ou endocrine. Les cellules pancréatiques exocrines sont organisées en acini alors que les endocrines sont en îlots. Ce pancréas accompagne les gros vaisseaux qui irriguent le foie. La vascularisation du foie est particulière : les capillaire sont différents des sinusoïdes (mais les sinusoïdes sont dans le foie).
On a donc un « hépato-pancréas » mais qui ne correspond pas à celui des invertébrés (mélange de deux types cellulaires).
B Le foie chez les batraciens.
Chez les amphibiens le foie présente une particularité. Il forme des lobes (2 ou 3). Chaque lobe est délimité à sa périphérie par du tissu lymphoïde accompagné de cellules pigmentaires. La partie interne des lobes contient des hépatocytes. Comme chez les poissons, ces hépatocytes sont organisés en travées anastomosées.
C Le foie chez les reptiles et les oiseaux.
Chez les reptiles et les oiseaux, le foie ressemble à celui des mammifères.
Le Rein/Appareil Urinaire.
I Généralités.
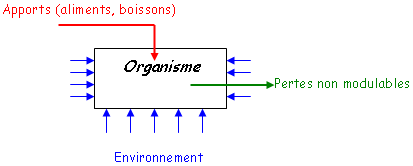
A Régulation.
Un organisme, par ses apports alimentaires, son environnement, par les pertes non modulables (eau pendant la transpiration) va devoir développer un système de régulation de son milieu intérieur : l’homéostasie.
1 Homéostasie hydro-électrolytique.
Le rein joue un rôle dans la régulation du capital hydrique, minéral et dans le maintien de la stabilité du pH dans le sang et le milieu intérieur.
2 Evacuation des déchets cataboliques.
Parmi les déchets du catabolisme, on trouve les déchets azotés comme NH3 (ammoniac) qui est toxique. Il va y avoir élimination de NH4+ (ammonium) ou d’urée ou d’acide urique. Cette régulation reflète la fonction exocrine du rein.
3 Régulation de la pression artérielle.
Le rein joue aussi un rôle endocrine, comme avec l’EPO ou le système rénine/angiotensine pour la régulation de la pression artérielle.
B Le rein de vertébrés.
Le rein est un organe massif, organisé de très nombreuses unités fonctionnelles : le néphrons. Le néphron est constitué de deux parties.
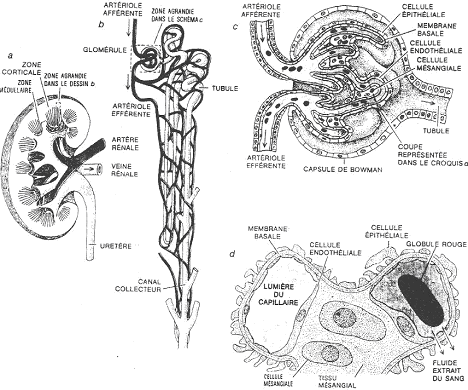
1 Le néphron.
a Le corpuscule.
Le corpuscule est l’endroit où se forme l’urine primitive, par filtration du sang. Le corpuscule est le glomérule accolé à la capsule. Le glomérule est une pelote de capillaires.
Le sang arrive par l’artère rénale et va être ultra-filtré. Le sang ressort par une veine efférente.
La vasa recta est un système circulatoire à contre courant.
b Les tubes.
Les tubes ont un trajet contourné. Ils ont pour fonction de conduire l’urine de la capsule jusqu’à l’extérieur (parfois, il n’y a pas de vessie).
Les tubes vont modifier l’urine en fonction du type cellulaire présent.
c L’organisation du néphron.
L’organisation du néphron est presque toujours la même. Les groupes les moins évolués ont les néphrons les moins évolués.
Le néphron ouvert est un glomérule pendu dans la cavité cœlomique, à son contact direct. Le sang ultra-filtré et l’urine transitent par la cavité cœlomique puis sont récupérés par un canal, le néphrostome, à épithélium simple, cuboïdal, cilié. On trouve cette organisation chez les alevins de poissons ou chez les larves de certains amphibiens.
Au cours de l’évolution, les néphrons se sont fermés. L’urine passe directement dans des tubes : ce sont les néphrons fermés. C’est le cas le plus général, sauf chez certains poissons marins qui ont des néphrons aglomérulés.
Dans ce cas, on n’a uniquement des tubes où se font les échanges, modifiant l’urine éliminée par un canal collecteur. è C’est une adaptation à des conditions de vie en forte osmolarité (à problème de la rétention d’eau).
Le néphron est toujours organisé en deux parties mais la subdivision des tubes peut être différente :
- Chez les non mammifères et non oiseaux, on a un corpuscule, un tube contourné proximal, un tube contourné distal et un canal de Wolf (évacuateur).
- Chez les mammifères et oiseaux, on a le corpuscule, le tube contourné proximal, le segment intermédiaire (anse de Henlé chez les mammifères), le tube contourné distal et des canaux collecteurs qui font déboucher l’urine dans un uretère secondaire. Il y a mise en place d’un canal spécifique à l’évacuation de l’urine.
L’apparition de ce segment intermédiaire régionalise le rein : on distingue maintenant une région corticale (le cortex) qui contient les corpuscules, et, une région médullaire (la médulla) qui regroupent les tubes contournés distaux et les anses de Henlé.
2 Le développement embryonnaire.
Le rein se met en place à partir d’une ébauche métamérisée. Cette disposition segmentaire implique un faible nombre de segments. La différenciation est progressive, d’avant en arrière et par étapes.
Il va y avoir mise en place d’un pronéphros (le rein définitif de poisson et des amphibiens). La disposition segmentaire va disparaître. Le nombre de néphrons augmente et l’urine est évacuée par le canal de Wolf (urine et produits sexuels).
Cette structure est temporaire chez les amniotes.
Il y a apparition du métanéphros qui voit se former un uretère secondaire spécialisé.
II Le rein de mammifères.
Les néphrons sont fermés, glomérulés et régionalisés. La circulation des vaisseaux sanguins souligne la ligne de séparation entre le cortex et la médullaire.
Il existe deux types de néphrons : les néphrons corticaux et les néphrons juxta-glomérulaires. Le fonctionnement de ces deux types est différent. Une différence morphologique importante est la différence de la taille de la anse de Henlé.
A Le corpuscule.
Le glomérule est contenu dans la dépression d’une capsule. En coupe, on voit dans le corpuscule, de nombreux capillaires sanguins : le glomérule. La capsule est constituée de deux feuillets : le feuillet pariétal (est bordé par un épithélium simple, squameux) et le feuillet viscéral (à épithélium formé de podocytes).
Les podocytes sont caractérisés par leurs prolongements cytoplasmiques se ramifiant, appelés, les pédicelles. Ces derniers s’appliquent étroitement autour des capillaires des glomérules.
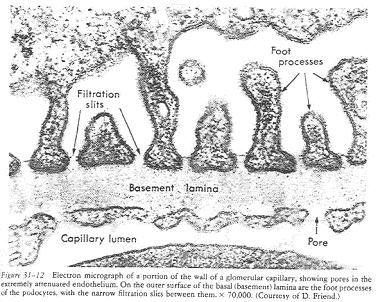
Entre les pédicelles et l’endothélium (des capillaires), on a les deux lames basales qui ont fusionné. Le sang arrive dans les capillaires sanguins, traverse cette barrière (de filtration) constituée d’un endothélium (fenêtré), d’une lame basale et d’un pédicelle.
L’espace ménagé entre les pédicelles s’appelle les « fentes de filtration ». La filtration du sang est un mécanisme passif se faisant grâce à la pression sanguine existant dans les capillaires. La pression dans les glomérule est forte (» pression aortique). Une diminution de cette pression entraîne une diminution de la filtration et une diminution de la quantité de l’urine.
La pression provoque le passage du sang vers l’espace de Bowman. Seul le plasma passe dans l’espace de Bowman (sans les grosses protéines).
Le tamis moléculaire vrai est la lame basale. Certains diabètes rénaux montrent une anomalie de cette lame basale. Au niveau des pores de l’endothélium, on observe un diaphragme (glycocalix anionique). Ils ne servent qu’à ralentir le passage des molécules.
Un autre type cellulaire est présent : ce sont les cellules mésengiales. Ce sont l’équivalent des macrophages et elles ont pour rôle principal, la dégradation des constituants de la lame basale. Elles permettent ainsi le renouvellement des constituants de cette lame basale. Le renouvellement est assuré par les podocytes.
Les cellules mésengiales servent aussi à l’élimination des grosses protéines (Ig) coincées dans la lame basale. On pense que ces cellules ont des caractéristiques communes aux péricytes (propriété contractile pour réguler le flux sanguin).
B Les tubes urinaires.
Les tubes urinaires sont les tubes contournés proximaux et les tubes contournés distaux. Ils conduisent l’urine mais vont aussi la modifier. Trois mécanismes entrent en jeu pour la modification : la diffusion, la sécrétion tubulaire, la réabsorption spécifique.
Les sécrétions sont les produits synthétisés par les cellules bordantes. Chaque tube a des caractéristiques reflétant ses cellules bordantes.
1 Le tube contourné proximal.
Le tube contourné proximal est bordé par un épithélium simple (palissadique ou cuboïdal), avec une bordure en brosse. Ces microvillosités montrent une augmentation de la surface d’échange. Ce tube peut aussi faire de l’endocytose (99% des acides aminés sont réabsorbés par ce tronçon ainsi que 100% des sucres).
Les cellules sont caractérisées par les microvillosités, des vésicules d’endocytose et au pôle basal, de nombreux replis membranaires (avec des mitochondries en alignement).
2 La anse de Henlé ou segment intermédiaire.
On trouve ces parties dans la médullaire.
La taille de ces anses varie en fonction de l’habitat de l’animal. Elle est courte quand l’animal vit près de l’eau et plus longue s’il vit dans des régions plus arides. Elle permet une lutte contre la déshydratation.
La anse de Henlé joue un rôle dans la concentration de l’urine.
La branche descendante est perméable à l’eau et aux sels. La branche ascendante est imperméable à l’eau et peut faire sortir activement les sels.
Si des sels sont dans le milieu interstitiel, ils entrent dans la branche descendante, donc, de l’eau sort. En descendant, l’urine va se concentrer. Elle se diluera en remontant, sous les mouvements d’eau : il se met en place un gradient cortico-médullaire. La concentration du tissu interstitiel s’accroît en allant vers le bas de la anse.
A la suite de cette anse, on trouve le tube contourné distal et le tube collecteur ; ils sont perméables à l’eau à certaines conditions. Comme le milieu interstitiel est concentré, le tube collecteur perd de l’eau et donc, concentre l’urine.
La anse de Henlé ne fait qu’augmenter la pression osmotique du tissu interstitiel pour que le tube collecteur perde beaucoup d’eau è économie d’eau. Il faut de l’ADH pour que le système fonctionne. Comme le tube collecteur est perméable à l’urée, il y a augmentation de la pression osmotique du tissu interstitiel (accroissement du phénomène).
La branche descendante diminue de diamètre (devient grêle) : il y a une surpression hydrostatique qui provoque la sortie d’eau.
La vasa recta est un système qui récupère l’eau qui sort des tubes : elle maintient le gradient.
La anse de Henlé, chez les mammifères, est bordée par un épithélium simple, squameux.
3 Le tube contourné distal.
Le tube contourné distal est bordé d’un épithélium simple, sans microvillosité, ni entérocyte, ni aldostérone. Il présente quelques replis membranaires avec alignements de mitochondries.
Le tube est capable de former la macula densa.
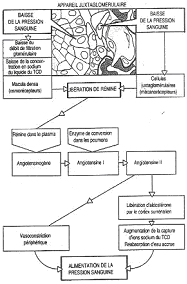
Les cellules du tube contourné distal sont capables, pour certaines, de se différencier en macula densa. Ces cellules deviennent des chémorécepteurs (sensibles à la concentration de sodium dans le tube contourné distal). La macula densa est au contact des artérioles afférentes et efférentes.
Certaines cellules musculaires lisses, autour de l’artériole afférente, se différencient en cellules à rénine. Ces cellules sont des barorécepteurs.
C’est l’appareil juxta-glomérulaire.
Quand la pression sanguine diminue dans l’artériole, ces cellules libèrent de la pro-rénine dans le sang. Elle va être clivée et donnera de la rénine.
Pour l’interaction avec l’angiotensine, voir cours de BGU03 (endocrinologie).
Une diminution de la concentration en sodium dans le tube contourné distal stimule la macula densa et stimule les cellules à rénine.
4 Le tube collecteur, l’uretère et la vessie.
Le tube collecteur a un épithélium simple, palissadique, sans bordure en brosse, sensible à l’ADH (réabsorption d’eau). Il permet un ajustement final de l’urine (concentration, volume).
L’urine passe dans l’uretère bordé d’un épithélium de transition (stratifié). Le même type d’épithélium est présent dans la vessie. Les cellules, au pôle apical, on une région de membrane plasmique semblant rigide : la membrane plasmique apicale possède des phospholipides avec beaucoup de cholestérol, ce qui donne cette rigidité membranaire. à Les régions rectilignes sont séparées par des régions plus souples.
Dans le cytoplasme, on a une mise en réserve de membrane plasmique (sous forme d’aiguilles à cause de la rigidité) qui pourra être réutilisée au pôle apical. C’est une adaptation des surfaces membranaires.
La vessie vide montre un épithélium épais alors que pleine, il semble fin. C’est une adaptation pour l’élasticité de la vessie.
III Rein et appareil excréteur d’oiseau.
Les néphrons sont fermés, glomérulés et régionalisés.
Le tube contourné proximal est comme celui des mammifères, ainsi que le tube contourné distal. Le segment intermédiaire est différent : l’épithélium est simple, cuboïdal. Les tubes collecteurs ont un épithélium palissadique.
Chez les oiseaux ont a des paquets de médullaire mélangés à du cortex.
On trouve un uretère (secondaire) mais pas toujours de vessie. Cet uretère est bordé d’un épithélium pseudostratifié, très haut, riche en cellules à mucus, avec souvent, du tissu lymphoïde.
Il y a une lutte contre la déshydratation par les phanères avec les plumes. Les oiseaux ont une urine riche en acide urique qui peut être évacué dans un faible volume d’eau : l’urine est quasiment solide.
IV Rein de batraciens.
Le rein de batraciens a des néphrons fermés, glomérulés et alignés (pas de segment intermédiaire). On voit souvent deux lignes de néphrons : on peut reconnaître les tubes contournés proximaux et tubes collecteurs.
Les tubes contournés proximaux et les tubes contournés distaux sont similaires à ceux des mammifères.
Le pôle urinaire est cilié.
V Rein de reptiles.
Le rein de reptile est une association de rein et de tissus génital. Les néphrons sont classiques ainsi que les tubes contournés proximaux, les tubes contournés distaux et les tubes collecteurs.
VI Rein de poisson.
Le rein de poisson occupe presque toute la longueur de l’animal (dans la cavité abdominale).
On trouve un rein céphalique (sans fonction urinaire) qui est un organe lymphoïde, et un rein postérieur, à fonction urinaire.
Les néphrons sont fermés, glomérulés et dispersés. Les tubes contournés proximaux sont classiques. Les tubes contournés distaux et les tubes collecteurs ont un épithélium simple cuboïdal.
Il n’y a pas de régionalisation du rein, ni d’alignement. On ne peut pas faire la distinction entre les tubes contournés distaux et les tubes collecteurs.
Le canal de Wolf ou uretère primitif a un épithélium pseudostratifié avec des cellules à mucus. Les corpuscules et les tubes baignent dans du tissu hématopoïétique. On voit aussi des cellules équivalent à des macrophages (tissu atrocytaire).
On trouve souvent une glande régulant la calcémie (corpuscule de Stanius) ainsi que des follicules thyroïdiens fonctionnels.
Le rein de poisson est un organe composite, comprenant différents types cellulaires et assurant diverses fonctions. Il ne présente pas de régionalisation.
L’appareil respiratoire.
I Généralités.
La respiration est l’ensemble des processus qui assurent l’oxydation de substrats organiques dont le but est de fournir de l’énergie.
Chez les poissons, on étudiera les spécialisations des échanges entre les branchies et le milieu extérieur.
A Demande permanente en O2.
La demande en O2 est permanente pour tous les animaux. Il existe peu de systèmes de stockage d’énergie. Chez certains insectes, on trouve des cellules géantes à hémoglobine. Les mammifères marins possèdent des muscles riches en myoglobine.
L’entrée d’O2 se fait toujours par diffusion passive. Il existe toutefois, des organes spécialisés. Les unicellulaires et les organismes de petite taille acquièrent l’O2 par diffusion au travers de la membrane, du tégument ou des tissus. Les animaux de plus grande taille doivent réduire les distances de diffusion. Il va donc apparaître une circulation au niveau de l’appareil respiratoire afin de transporter l’O2 venant de l’extérieur jusqu’aux cellules.
Dans le cas de la respiration tégumentaire, le sang circule juste sous le tégument que l’O2 traverse sans difficulté.
Afin d’augmenter le rendement de la respiration, il va y avoir un accroissement des surfaces d’échange (invagination ou évagination). L’évagination est le système de la branchie. Les poumons viennent d’une invagination.
La diffusion simple est le seul mécanisme de passage de l’O2.
B Le transport de l’O2.
Le pigment circulant de l’O2 est l’hémoglobine. Le pigment non-circulant de cette molécule est la myoglobine.
Une fois l’O2 diffusé, il se fait capter par les pigments. Dans le cas de l’hémoglobine (métallo-protéine), il y a nécessité d’un ion (Fe2+, Cu2+) pour capter la molécule. Le fer ferreux associé à une protoporphyrine forme un hème. Ce dernier est tétramérique chez les vertébrés supérieurs. Les pigments subissent un changement de conformation en fonction de la saturation en O2.
L’hémoglobine a une affinité différente pour l’O2 en fonction des espèces et des conditions de vie.
La myoglobine, pigment réservoir, est présente dans les muscles, en grande quantité chez les mammifères marins.
C Le transport du CO2.
Le CO2 est relativement soluble : 7% est dissout. Majoritairement, on trouve la forme H2CO3 :
CO2 + H2O ó H2CO3 ó HCO3- + H+ (70%).
Il existe aussi une liaison du CO2 à l’hémoglobine, mais sur des sites différents de ceux à O2. Hb + CO2 = carbamine.
R-NH2 + CO2 ó R-NHCOO- + H+
II Les divers modes de respiration.
A La respiration tégumentaire.
Cette respiration existe chez les invertébrés, les amphibiens et les reptiles et nécessite une forte vascularisation sous-cutanée. Dans certains cas, on trouve des replis cutanés très nombreux avec parfois des papilles dermiques qui permettent l’augmentation de la surface d’échange.
Cette respiration est importante chez les reptiles. Chez les batraciens, on peut trouver trois types de respiration.
B La respiration branchiale.
Les surfaces spécialisées viennent de l’adaptation de l’épithélium pharyngé. Il y a diffusion épithéliale avec le milieu extérieur par le gradient de pressions partielles.
· La convection circulatoire interne : il existe deux types de sang : un sang oxygéné (pauvre en CO2) issue des appareils respiratoires et un sang veineux pauvre en O2 (riche en CO2).
· La diffusion tissulaire : il y a des échanges entre les tissus et les capillaires par diffusion simple.
· La ventilation : le débit d’air et d’eau qui transite dans les cavités respiratoires provient de la mécanique ventilatoire.
La source d’O2 (dissout) est l’eau. Cette respiration branchiale concerne les vertébrés aquatiques inférieurs et les larves d’amphibiens.
La ventilation se fait par un courant d’eau entrant par la bouche et qui ressort par les fentes operculaires : la respiration branchiale est unidirectionnelle et ne contient pas d’espaces morts anatomiques.
La mécanique ventilatoire :
- S’il y a une double pompe qui fonctionne (une pompe buccale et une pompe operculaire), il va se créer un gradient de pression et donc un courant d’eau unidirectionnel.
- S’il n’y a pas de double pompe, un déplacement permanent de l’animal, bouche ouverte (requins) est obligatoire.
L’appareil respiratoire présente :
- Un faible volume mais une grande surface de contact.
- Des branchies externes larvaires.
- Des branchies internes pharyngiennes : ce sont des replis épithéliaux très vascularisés et irrigués par des arcs aortiques.
En général, 4 à 5 paires de branchies sont accrochées à un arc branchial. Les branchies sont des alignements de lames (filaments) formant des feuillets. Ces lames portent des lamelles qui sont les échangeurs respiratoires. Une coupe de lame montre des lamelles de part et d’autre de la lame. La lamelle possède un épithélium simple squameux (ou épithélium respiratoire). L’O2 dissout dans l’eau traverse l’épithélium et se retrouve au contact de « capillaires » sanguins. A ce niveau, le sang circule dans des lacunes sanguines et non dans des capillaires. Ces lacunes sont portées par des cellules en pilastres : ce sont des prolongements cytoplasmiques. Les cellules en pilastres ont un rôle important car elles possèdent un cytosquelette particulier qui empêche la lamelle de se refermer (s’écraser) à cause de la pression d’eau. Ces cellules ont aussi des propriétés contractiles : elles règlent les diamètres des lacunes et modifient ainsi le débit sanguin.
L’épithélium basal (des lamelles) est stratifié et comporte des cellules à mucus. Ces cellules ont sûrement un rôle de protection mécanique.
On trouve aussi des cellules à chlorure qui ont un rôle important dans le maintien de l’osmolarité du poisson. Ces cellules sont caractéristiques des cellules ayant pour fonction l’excrétion ou l’absorption de molécules. Elles présentent aussi de nombreux replis membranaires à la base des cellules. Entre elles, ces cellules sont attachées par des jonctions fermées. On voit aussi apparaître des cellules accessoires établissant des jonctions lâches avec les cellules à chlorure. C’est une modification de l’épithélium et donc, une modification de fonctionnement.
Au niveau des branchies, on trouve des cellules à rôle de régulateur de l’osmolarité. Le sel entraîne des risques de déshydratation compensés par la synthèse d’une urine très concentrée et peu abondante. Les cellules auxiliaires viennent modifier les cellules à sel.
En eau douce, la tendance est différente : les animaux boivent peu et les entérocytes absorbent le sel. L’urine est abondante. Les branchies ont un rôle respiratoire mais aussi un excréteur/osmorécepteur.
C La respiration pulmonaire aérienne.
Cet appareil respiratoire a pour origine la région antérieure du tube digestif. La ventilation se fait par des mouvements de va-et-vient (inspiration et expiration).
Chez les oiseaux, la circulation d’air est réalisée par l’intermédiaire de sacs.
1 Les amphibiens.
Chez les amphibiens, les poumons sont sacculaires. Ils sont cloisonnés afin que ces cloisons accroissent la surface d’échange. On distingue des cloisons primaires et des cloisons secondaires. L’épithélium est simple, squameux.
La mécanique ventilatoire se fait souvent par soulèvement du plancher buccal.
2 Les reptiles.
Les poumons reptiliens sont de tous les types possibles : du plus simple (comme les amphibiens) aux plus complexes (ressemblant à ceux des mammifères).
Au sommet des cloisons, on trouve des masses de muscles lisses : les septes, à épithélium cuboïdal. C’est l’épithélium des cloisons qui est respiratoire.
3 Les oiseaux.
L’appareil respiratoire des oiseaux est modifié pour une meilleure adaptation au vol. On remarque que la chauve-souris possède des caractéristiques identiques à celles des oiseaux.
Ici, la ventilation est complexe. Les poumons sont compacts, quasiment rigides. Il n’y a pas de mouvement de cage thoracique. Ce sont cinq paires de sacs aériens qui jouent le rôle de soufflet.
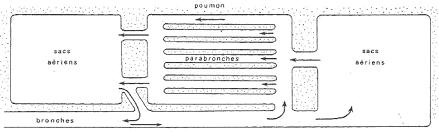
La circulation de l’air.
Il faut deux cycles d’inspiration et expiration pour que l’air passe dans tout l’appareil respiratoire et en ressorte. Les sacs jouent un rôle de soufflets qui maintiennent dans les conduits aériens du poumon, une circulation continue et unidirectionnelle.
La première inspiration fait passer l’air (extérieur) au premier sac (postérieur). La première expiration fait passer l’air de ce sac jusqu’aux poumons. La seconde inspiration fait passer l’air des poumons au sac antérieur et la seconde expiration amène l’air du sac antérieur à l’extérieur du corps.
Cette mécanique originale permet de toujours avoir de l’O2 disponible.
L’air passe du sac postérieur à la bronche qui se divise en parabronches. L’air y circule en traversant des capillaires aériens. Ces derniers sont accolés à des capillaires sanguins, ce qui favorise les échanges.
La disposition des capillaires aériens et sanguins est telle qu’il s’établit un courant croisé (un « cross current »). Ce système est très proche de système de contre-courant de la branchie : il y a accroissement de l’extraction de l’O2.
Le gradient d’oxygène qui est créé est toujours favorable au passage de l’O2 du milieu extérieur vers le sang. è On a une grande efficacité du système respiratoire.
4 Les mammifères.
Il n’existe pas de courant croisé ni inversé chez les mammifères. L’O2 va au fond d’une alvéole et arrive au contact de l’épithélium, lui-même au contact des capillaires sanguins.
L’appareil respiratoire.
L’intérieur du nez est constitué d’un épithélium stratifié cilié, spécialisé pour l’olfaction. Des poils vont filtrer l’air. Cet air va s’humidifier et se réchauffer.
a Le pharynx.
Le pharynx est le lieu de jonction des conduits aérien et digestif.
b Le larynx.
Le larynx est dans la continuité du pharynx. C’est un organe développé et modifié en vue de la phonation. Il y a présence de replis fibro-musculaires qui donnent les cordes vocales. Le larynx possède une charpente cartilagineuse.
L’épithélium est stratifié squameux. Dans le bas larynx, l’épithélium est de type trachée (pseudostratifié, cilié, riche en cellules à mucus).
c La trachée.
La trachée est un tube maintenu béant par des anneaux cartilagineux. L’épithélium est pseudostratifié, cilié, riche en cellules à mucus.
Sous l’épithélium, on trouve des glandes trachéales synthétisant du mucus servant à colmater les particules et les poussières inspirées. Les cellules ciliées vont faire remonter ces particules afin de les expectorer. La trachée se divise en bronches.
d Les bronches.
Les bronches sont caractérisées par des plaques cartilagineuses. L’épithélium est simple pseudo-stratifié. L’épithélium intra-pulmonaire est simple cuboïdal.
Il existe des glandes bronchiques dans le tissu conjonctif. Elles donnent aussi du mucus servant à expectorer les particules inspirées.
e Les bronchioles.
Les bronchioles ne présentent ni glandes, ni cartilage. Elles sont constituées d’un épithélium simple avec des cellules ciliées et de rares cellules à mucus. On trouve aussi les cellules de Clara (non ciliées). Ces cellules synthétisent du lysozyme, grains de sécrétions anti-bactériens.
Autour des bronchioles (comme pour les bronches), on trouve du muscle lisse en grande quantité. A la fin de l’arbre respiratoire, l’air va dans l’atrium pour être déversé dans des sacs alvéolaires puis au fond des alvéoles.
Les capillaires présents sont vrais (à la différence des branchies).
En microscopie électronique, on voit que la barrière air/sang est constituée d’un épithélium simple, squameux (avec lame basale) et d’un endothélium de capillaires sanguins (avec sa lame basale). Parfois, les deux lames fusionnent.
Chez les mammifères, on trouve deux types cellulaires :
- Les pneumocytes I. Ce sont des cellules qui émettent de longs prolongements cytoplasmiques, très fins. Ils coexistent avec l’autre population cellulaire.
- Les pneumocytes II. Ils font aussi parti de l’épithélium où ils établissent des jonctions. Leur principale fonction est la synthèse de surfactant (phospholipides), pouvant être déversés à la surface des alvéoles et empêcher le collapsus des alvéoles.
Remarque : chez les batraciens, il n’existe qu’un type de pneumocytes qui joue les deux rôles.
L’intérieur des alvéoles montre des macrophages alvéolaires (dans la lumière) qui participent à la défense de l’organisme. Il existe aussi des macrophages entre les capillaires sanguins.
Tout le long de l’appareil respiratoire, il existe des amas lymphoïdes utiles à la défense immunitaire.
L’appareil circulatoire.
I Les capillaires.
Les plus simples capillaires sont les sinusoïdes, délimités seulement par les cellules endothéliales formant un épithélium simple, pénétré de pores permettant des échanges macromoléculaires entre le sang et le tissu irrigué (pas de lame basale). « Endothélium » s’applique au trajet du sang. Les sinusoïdes sont soumis à la pression du sang circulant. Ils sont typiques du foie.
La plupart des capillaire sanguins ont une paroi plus complexe que celle des sinusoïdes : ce sont des capillaires à endothélium continu ou discontinu. Dans le cas d’un épithélium discontinu, il existe entre les cellules endothéliales, des jonctions lâches permettant le passage de globules blancs (dans la rate). Généralement, l’endothélium continu est formé de cellules jointives. Il peut être fenêtré ou non : des pores permettent le passage de macromolécules (dans le rein). S’il est continu, on est dans le pancréas. Ils ont une lame basale (sauf les sinusoïdes). Il peut y avoir présence ou non de péricytes à la périphérie de la lame basale. Les péricytes ne forment jamais de couche continue mais seulement des sections où le corps cellulaire s’étend à la périphérie des capillaires.
Les cellules endothéliales présentent de petites vésicules connectées aux membranes ou au cytoplasme. Elles permettent le transport de macromolécules vers la face opposée. La libération se fait par exocytose (deux directions) è c’est la transcytose.
Il existe une exception à ce phénomène : les cellules endothéliales du cerveau. Aucune endocytose n’est visible. Les capillaires sont des barrières pour les échanges de macromolécules. Ils apportent les éléments nutritifs mais seulement par des petites molécules. Il y a protection des cellules nerveuses.
II Les petites veines ou artères.
La paroi des vaisseaux devient plus étoffée. On trouve trois tuniques.
A L’intima.
L’intima est une lame élastique. Elle est au centre du vaisseau, constituée de cellules endothéliales reposant sur une lame basale. Dans la lame basale, il existe des fibres d’élastine qui constituent la lame élastique interne.
B La média.
La média est constituée de cellules musculaires lisses mêlées à des fibres élastiques (élastines).
C L’adventice.
L’adventice est une tunique comprenant des capillaires sanguins et des filets nerveux. Elle contient aussi du tissu conjonctif. A l’interface entre média et adventice se trouve une lame élastique externe.
III Grosses artères et grosses veines.
A Grosses artère : l’aorte.
La média est particulièrement riche en fibres d’élastine avec peu de fibres musculaires : il va y avoir amortissement du flux sanguin à la sortie du cœur : l’aorte, c’est une artère élastique.
B Grosse veine.
Les grosses veines sont composées de trois tuniques, comme les artères. Seules les proportions des tuniques vont varier. Pour des vaisseaux de même calibre, on observe :
Les tissus endocrines.
Les glandes endocrines peuvent être classées en deux catégories :
- Les glandes synthétisant des hormones peptidiques
- Les glandes synthétisant de petites molécules, souvent hydrophobes, comme les hormones stéroïdes.
I Les glandes à hormones stéroïdes.
A La glande médullosurrénale.
La glande surrénale est formée, chez les mammifères, de deux tissus d’origine embryologique distincte :
- Le tissus inter-rénal qui dérive de l’épithélium de la cavité cœlomique de l’embryon (mésoderme).
- Le tissu chromafine qui a pour origine l’ectoderme des crêtes neurales.
Ces deux tissus, chez les vertébrés, sont présents sous forme de glandes rapprochées. Chez les mammifères, on parle de glande surrénale.
La partie d’origine mésodermique forme le cortex et la partie d’origine ectodermique donne la médullaire.
Chez les vertébrés non mammifères, ces tissus ne forment pas forcément une glande. Le poisson a du tissu inter-rénal dans la partie antérieure de la cavité abdominale, en avant du rein à fonction excrétrice.

Chez les amphibiens, le rein a une forme allongée et le tissu inter-rénal est placé à la surface du rein, sur toute la longueur de l’organe. La partie de tissu chromafine peut occuper une région différente, et ne pas être associée à l’inter-rénal.
C’est le tissu inter-rénal qui donne les hormones stéroïdes.
La surrénal, chez les mammifères, est constituée de cordons cellulaires formés de cellules de grande taille. On observe trois zones : la zone glomérulée où les cellules sont organisées en mimant l’allure d’un glomérule ; une zone fasciculée qui est la plus étendue ; une zone réticulée où les cellules sont petites, positionnées en cordons anastomosés.
Les cellules de taille importante ont un noyau central, sphérique ; l’aspect du cytoplasme est spongieux, à cause des gouttelettes lipidiques.
En microscopie, quelle que soit la région de prélèvement, l’aspect cellulaire est identique. Le réticulum endoplasmique lisse est particulièrement développé. On observe aussi de nombreuses mitochondries. Le golgi peut être étendu. Il y a peu de réticulum endoplasmique rugueux.
Les stéroïdes dérivent des lipides (acétate). Ils passent dans une chaîne de synthèse qui conduit, au niveau des membranes de réticulum endoplasmique lisse, à du cholestérol. Du cholestérol à la prégnénolone, il y a de nombreuses étapes qui se déroulent dans la mitochondrie. Il y a intervention des enzymes mitochondriales. La prégnénolone est le précurseur commun de beaucoup d’hormones stéroïdes. Cette molécule est aussi produite par d’autres glandes endocrines.
Au final, on obtient des glucocorticoïdes, de l’aldostérone.
Les étapes conduisant à ces stéroïdes se déroulent dans le réticulum endoplasmique lisse.
Tout ceci est aussi vrai pour :
- Les cellules produisant la testostérone (cellules de Leydig) formant le tissu interstitiel des testicules.
- Le corps jaune qui synthétise des œstrogènes.
Il y a une homologie de structure.
On ne trouve jamais de grains de sécrétions car ces petites molécules ne sont pas accumulables. Elles sont hydrophobes et sont sécrétées immédiatement après leur production.
B La thyroïde.
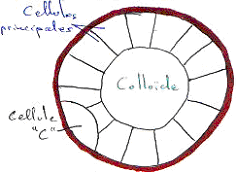
La thyroïde ne donne que des hormones de petite taille (T3 et T4). Elles sont des dérivés de la tyrosine et portent des groupements iodés. Elles sont susceptibles, par leur nature, de traverser les membranes plasmiques.
Les follicules thyroïdiens sont des sphères closes qui, en coupe, apparaissent constitués d’un épithélium simple, cuboïdal. Une cavité centrale est délimitée et la lame basale est autour de chaque follicule (en périphérie).
Les follicules présentent deux catégories cellulaires :
- Les cellules principales. Ce sont les plus nombreuses. Elles sont caractérisées par leur capacité à concentrer l’iode et au fait qu’elles exposent leur pôle apical vers la lumière.
- Les cellules « C ». Ce sont les cellules productrices de calcitonine. Elles sont caractérisés par leur non-concentration d’iode et à la présence, dans leur cytoplasme, de grains de sécrétion.
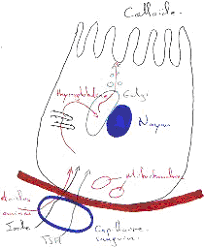
Les cellules principales ne synthétisent pas directement les hormones thyroïdiennes. Elles forment d’abord une glycoprotéine de grande taille (660KDa), la thyroglobuline. Celle-ci est ensuite déversée dans la colloïde (en T3 ou T4). L’hormone thyroïdienne pourra ensuite être produite. è Il y a deux phases dans la production : la première consiste à produire un précurseur de grande taille qui sera utilisé pour produire les hormones finales.
L’organisation de ces cellules est différente de l’organisation productrices de stéroïdes.
1 Les cellules principales.
A leur pôle apical, on trouve des microvillosités dispersées ; le pôle basal est appliqué contre la lame basale. On trouve de nombreux capillaires sanguins dans le tissu conjonctif voisin.
De nombreuses glycosylations sont faites dans le golgi. Ensuite, de petites vésicules se forment et amènent cette glycoprotéine vers la membrane plasmique apicale. Au niveau de cette membrane, il va y avoir fixation d’un iode sur le motif tyrosine de la protéine (cf. BGU 03). C’est sous cette forme qu’est mise en place la colloïde.
S’il n’y pas de stimulation par la TSH, tout s’arrête.
Quand il y a stimulation, les cellules deviennent capables de phagocyter (par des pseudopodes), la thyroglobuline iodée (TGI). Il y a apparition de grosses boules dans la face apicale.
Après l’internalisation de TGI, il va y avoir digestion intra-cellulaire avec des lysosomes primaires qui donnent des lysosomes secondaires (réactifs + enzymes). Il y a lyse de la chaîne peptidique et libération de l’hormone thyroïdienne. Cette dernière molécule va diffuser à travers la membrane plasmique afin d’aller dans le sang.
Ces cellules ont un réticulum endoplasmique rugueux bien développé, un golgi actif et excrètent dans la colloïde, stockent et sécrètent.
2 Les cellules « C ».
Les cellules « C » ont un aspect différents des cellules principales. Elles synthétisent la calcitonine, peptide de 32 acides aminés. Une fois produite, elle passe dans des grains de sécrétion et sera déversée dans le sang par exocytose.
Chez les poissons, batraciens, reptiles, oiseaux, les cellules « C » forment le « corps ultimo-branchial », séparé de la thyroïde mais situé dans son voisinage, ventralement, en avant du corps. Les poissons ont aussi de la thyroïde dans le rein, sous forme de follicules.
II Les glandes à hormones peptidiques.
A coté de la thyroïde, on trouve les parathyroïdes. Elles sont accolées à la thyroïde. Il n’y que chez le poisson où leur équivalent, le corpuscule de Stanius, est associé au rein.
Les parathyroïdes sont formées par des cellules à aspect hétérogène. On trouve trois types cellulaires :
- Les cellules principales. Elles sont actives dans la production de PTH.
- Les cellules oxyphiles.
- Les cellules claires.
Il semble qu’en fait, toutes ces cellules soient productrices de PTH et les différents aspects seraient des états fonctionnels distincts.
La PTH a un rôle hypercalcémiant : elle favorise la mobilisation du calcium à partir des os et limite les pertes à partir des reins.
L’hormone produite par le corpuscule de Stanius a un rôle hypocalcémiant et a pour cible les branchies.
III Glandes endocrines comme l’hypophyse ou le pancréas.
A L’hypophyse.
| L’hypophyse vient de l’union de deux tissus d’origine embryonnaire différente. Un tissu vient de l’endoderme, l’autre vient du neuroectoderme. |
|
Un système vasculaire porte permet le transport des molécules produites par l’hypothalamus, jusqu’à l’hypophyse.
Depuis longtemps, on sait que l’hypophyse contient de nombreuses catégories cellulaires. On reconnaissait trois types :
- Les cellules acidophiles ; colorables par la fushine (rose) ou par l’orange G (orange).
- Les cellules basophiles ; colorables par les colorants nucléaires.
- Les cellules chromophobes ; non colorables.
Maintenant, on distingue les cellules acidophiles, les cellules basophiles qui, repérées par le PAS, donnent deux types cellulaires : les cellules à grains de grande taille à cellules synthétisant des hormones en relation avec l’appareil génital (taille hétérogène), synthétisent l’hormone somatotrope (cellules à STH) ; les cellules à TSH.
Remarque : ce sont les grains que l’on regarde. En microscopie électronique, ces cellules sont identifiables par la taille, la forme et la densité aux électrons des grains de sécrétion. La TSH donne une tache noire. C’est grâce à l’immunocytologie que l’on a pu mettre en évidence les colorant et les fonctions.
Si un animal est soumis à un jeûne alimentaire prolongé, il va produire de la STH.
Les critères de coloration ne peuvent être utilisés que lorsque les cellules sont en un état de fonctionnement moyen.
On observera des décharges de grains que si la cellule (hypophysaire) est stimulée.
B Le pancréas endocrine.
Le pancréas endocrine représente 1% du pancréas. Ce sont les îlots de Langerhans. Les cellules ont un aspect hétérogène. On distingue quatre catégories cellulaires :


L’ensemble des cellules est organisé par rapport aux capillaires. Elles organisent entre elles des jonctions fermées. L’îlot montre des interactions entre les cellules le composant.
A côté de la fonction endocrine, on doit noter une fonction paracrine : les produits de synthèse ont une action sur les cellules voisines à la cellule productrice.
Les îlots montrent plus de cellules b que de cellules a. Les cellules PP sont minoritaires. Ceci n’est vrai que pour certains îlots. Chez le rat, il existe de fortes différences entre les îlots de la région dorsale du pancréas et les îlots de la partie ventrale.
Les proportions varient lors de pathologies. Les rats diabétiques montrent une répartition cellulaire différente de la normale.
Résultats obtenus par analyse se coupes sériées, analysées par plusieurs marqueurs immunofluorescents :

A. Morphologie
On ne voit rien au MO. Le hyaloplasme est vide photoniquement, il a un aspect hyalin (rien).
Il est très hétérogène au MET.
On observe des réserves (glycogène, globules lipidiques, cristaux protéiques). Ces réserves sont les éléments figurés.
On observe aussi des ribosomes libres (non associés à l'exomembrane), et le cytosquelette (éléments protéiques à structure filamenteuse, ils sont soit isolés, soit organisés en faisceaux).
Le hyaloplasme occupe tout l'espace entre les organites, il représente 50 % du volume cellulaire.
B. Biochimie
Le hyaloplasme est riche en eau. H2O représente 80 à 85 % de sa composition.
Le reste est une très grande variété de molécules : protéines solubles (enzymes) / protéines insolubles (structure) / lipides et précurseurs : acides gras, alcools / sucres et précurseurs : oses / acides nucléiques (en majorité des ARN, ARNm, ARNt, ARNr et nucléotides. / sels minéraux, ions (=> milieux tampon)
Ces éléments viennent du milieu extracellulaire. Ils passent la membrane en empruntant des transporteurs qui régulent les échanges, ou en empruntant des structures digestives par endocytose.
Physiquement, il s'agit d'un gel colloïdal à pH 6,8. Toutes les molécules qui le composent sont liées par des liaisons H et hydrophobes, liaisons faibles, labiles (qui peuvent se casser).
Si les liaisons sont nombreuses, les hyaloplasme a un aspect gel. Si les liaisons sont peu nombreuses, les hyaloplasme a un aspect sol.
C. Physiologie
Carrefour métabolique
Le cytosol est un carrefour métabolique : pratiquement tous les cycles ont leur origine dans le cytosol. Il a un rôle prépondérant dans l'initiation des réactions. En effet il contient tous les précurseurs des voies anaboliques et cataboliques. Ex pour la fabrication de l'ATP. Le premier processus, la glycolyse, s'effectue dans le hyaloplasme et aboutit à la formation d'acide pyruvique. L'ATP se forme dans la mitochondrie grâce au cycle au cycle de Krebs. A la fin, l'ATP, le CO2, et l'H2O résultants se retrouvent dans le hyaloplasme.
Génération de mouvement
C'est le lieu où l'on va générer des mouvements. (intracellulaires ou cellulaires)
Modifications post traductionelles
1) Phosphorylation réversible : l'activité de 10% des protéines est régulée par la phosphorylation / déphosphorylation. (kinases / phosphatases)
2) Glycosylation : des sucres sont accrochés sur les protéines (glycoprotéines). Seules les glycosylations de type "O" se font dans le hyaloplasme. (elles restent relativement simples). Ex : Nucléoporines, cytosquelette, facteurs protéiques de l'ARN pol II. Le mécanisme fait appel à l'enzyme glycosyl transférase qui ajoute au minimum un résidu N-acétyl-glycosamine sur les acides aminés Sérine ou Thréonine.
3) Modification par des acides gras <=> Acylation. Les protéines cytosoliques peuvent se lier au plasmalemme ou à des système d'endomembrane (RE, Golgi). Les protéines hydrophiles deviennent hydrophobes. Ex protéine G : trimérique ou monomérique que à activité GTPasique. Ex lamines : protéines qui peuvent s'associer à la membrane du noyau.
4) Protéolyse cytosolique : pour les protéines à demie vie courte. (protéines de régulation, enzymes). La séquence en a.a. N-ter est caractéristique : 12 a.a. sont déstabilisants parmis lesquels Isoleucine, Leucine, Arginine ; 8 a.a. stabilisent la protéine dont la Méthionine et la Glycine. L'ubiquitine (présente dans toutes nos cellules / 70 aa) s'accroche de manière covalent sur les Lysines des protéines et les guide vers les protéasomes pour y être dégradées. 30% des protéines sont détruites par cette voie. Le protéasome est un complexe multi enzymatique à activité protéasique fonctionnant à pH neutre alentour de 6,8 / 7. C'est un cylindre creux de 500 Ǻ de hauteur et 200 Ǻ de diamètre.
5) Adressage des protéines synthétisées : il existe des facteurs d'adressage qui reconnaissent les protéines grâce à certaines séquences spécifiques d'adressage. Ex la particule de reconnaissance de la séquence signal, SRP, adresse les protéines à l'intérieur du RE ou la protéine de liaison de la séquence de localisation nucléaire, NLS - BP.
Les protéines chaperonnes sont des protéines d'accompagnement pour d'autres protéines. Elles existent dans le hyaloplasme, les mitochondries et le RE. Ex : protéines de choc thermique, (Hsp) / protéines de stress. Ex : ubiquitine, celle ci est une très petite protéine. Toutes ces molécules sauf l'ubiquitine fixent l'ATP, l'ADP et ont une activité ATPasique. Cette activité est régulée par des protéines co-chaperons. Ce sont des molécules de constitution et/ou inductibles. Elles ont pour fonction d'adresser les protéines vers les mitochondries ou, comme la protéine BIP de transloquer des protéines à travers la membrane du RE, ou prennent en charge les protéines néoformées pour qu'elles acquièrent leur structure 3D. Ex : Hsp70 (70 kDa), s'accole aux protéines en cours de formation. Avec l'hydrolyse d'une molécule d'ATP, elle donne une nouvelle conformation à la protéine. De plus, Hsp70 empêche les agrégats de protéines. Hsp60 (60 kDa), quant à elle, se polymérise pour donner des structure cylindriques de 14 protéines. Elle nécessite pour fonctionner 14 molécules d'ATP.
A. Morphologie
Ils sont invisibles au MO
Sur coupe au MET on observe des organites limités par une membrane biologique classique.
Æ 0,5 µm jusqu'à exceptionnellement 1 à 2 µm
Le contenu de ces vésicules est uniformément opaque aux électrons.
Les lysosomes contiennent en abondance des protéines enzymatiques.
La face luminale (¹ cytosolique) est recouverte d'une couche glucidique qui protège la membrane.
B. Biochimie
Les lysosomes sont des sacs à enzymes. Il y a beaucoup d'hydrolases qui agissent de façon optimale à pH 5 ; ex : protéinases, lipases, glucosidases.
Membrane du lysosome
Elle est normalement imperméable aux macromolécules biologiques. Les enzymes présentes à l'intérieur sont capables de détruire n'importe quelle molécule mais ne peuvent s'échapper grâce à la couche glucique. Celle est constituée de la juxtaposition de domaines glycoprotéiques. Ce sont les protéines Lamp1 et Lamp2, associées à la membrane du lysosome, qui donnent naissance à la couche glucidique.
Une pompe à protons est chargée de maintenir un pH acide / en augmentant [H+]. Énergie dépendante, elle consomme le GTP. Plusieurs peptides constituent cette pompe à protons.
Dégradation
Les protéines sont dégradées en acides aminés, les glucides en oses, les acides nucléiques en nucléotides. Les produits de la dégradation peuvent traverser la membrane pour aller dans le cytosol. Des perméases permettent l'expulsion.
3 Types de lysosome
Lysosomes primaires / natifs : petite vésicule opaque aux électrons. Ils sont au repos et non fonctionnels.
Lysosomes secondaires : ceux-ci sont fonctionnels. Plus gros ils ont fusionnés avec une vésicule / vacuole d'endocytose.
Lysosomes tertiaires ou "corps résiduels" : résultat de l'hydrolase ils renferment les déchets non dégradables et sont ré expulsés en dehors de la cellule.
C. Physiologie
Phénomènes de digestion
Digestion intracellulaire : c'est leur fonction principale, c'est la fin de l'histoire des vésicules d'endocytose.
Digestion extracellulaire : elle ne s'effectue pas chez toutes les cellules, mais seulement chez les macrophages des tissus osseux : ostéoclastes et chondroclastes. Ici les hydrolases sont expulsées dans le milieu extracellulaire. Elles dégradent perpétuellement la matrice osseuse et cartilagineuse. Le renouvellement de la matrice étant assuré par les ostéoblastes et chondroblastes.
Autres rôles
Défense passive : Ex dégradation des bactéries. Mais un certain nombre de bactéries sont indigestes à cause d'une paroi chimiquement complexe, parmis lesquelles on compte le bacille de Koch et la lèpre.
Phénomènes sécrétoires : La thyroxine est synthétisée par la thyroïde sous forme de précurseur : la thyroglobuline qui une fois iodée est réabsorbée. La dégradation dans le lysosome est presque totale, seul reste la thyroxine.
Involution d'organe : Ex embryogenèse du rein, d'abord complexe, le rein partiellement détruit aboutit au rein final. L'autodestruction contrôlée des tissus est due à l'action des lysosomes, d'ailleurs appelés pour l'occasion autophagosomes.
D. Pathologie
Destruction non contrôlée des cellules
Phénomène différent de l'involution, due à l'accumulation dans les lysosomes d'élément organiques ou minéraux non dégradables. Les enzymes lytiques se répandent suite à la perforation de la membrane du lysosome.
Cas de composés organiques non digérés : ex l'acide urique : des cristaux d'urate de sodium s'accumulent dans les synoviales, les macrophages les bouffent mais malheureusement ils ne possèdent pas d'uratase. Les macrophages dégradés explosent ce qui déclenche le processus inflammatoire.
Cas de composés métaboliques :
Pneumoconiose : destruction des poumons à cause d'accumulation de silice - Silicose, accumulation d'amiante - Asbestose.
Maladie de Tay - Sachs
Maladie autosomique récessive dont le gène est situé sur le chromosome 15. Normalement l'enzyme hexosaminidase décompose le GM2, un glycolipide membranaire abondant dans le plasmalemme des cellules nerveuses. Dans les cellules atteintes, le GM2 n'es pas dégradé. Le renouvellement est déséquilibré, la concentration cellulaire en GM2 augmente entraînant un disfonctionnement des cellules nerveuses.
Mucoviscidose de type 2
Phénomène de thésaurisme, les métabolites s'accumulent dans le lysosome suite à une anomalie du transit (ex perméase déficiente). On assiste alors à un relargage des produits de dégradation.
Le plasmalemme est la structure qui définit la cellule. C'est un constituant universel de toutes les cellules vivantes.
Au MO, on observe une bordure plus ou moins réfringente colorée par des sels d'argents (Ag NO3). Le MET révèle une structure tripartite épaisse de 75 à 80 Ǻ constituée de deux feuillets opaques aux électrons de 25 Ǻ. Ces dimensions restent valables quelles que soient les cellules. L'espace intercellulaire est de 150 à 300 Ǻ. Pour les cellules épithéliales (en contact avec la lumière) on trouve généralement à la face apicale (¹ face basale) une structure appelée cell coat ou revêtement cellulaire ou encore glycocalix. C'est un ensemble de petites fibres posées perpendiculairement au feuillet externe qui réalisent une couche supplémentaire de 250 à 2500 Ǻ.
Les membranes biologiques sont un complexe lipo-protéique. Dans le cas du plasmalemme, il y a aussi des sucres, c'est un complexe glyco-lipo-protéique. Le rapport lipides-protéines du plasmalemme vaut un : R=P/L=1, c'est à dire que la masse de lipides et égale à la masse en protéines.
Leur PM est faible : » 1000 Da, ce qui fait qu'il y a à peu près 50 molécules de lipides pour une protéine. Les lipides sont des molécules polarisées, amphiphiles, avec un pôle hydrophobe (acide gras) et un pôle hydrophile (alcool). Dans toute membrane biologique, les lipides sont organisés sous forme d'une bicouche. Les pôles hydrophobes se faisant face, ils sont liés par une liaison hydrophobe.
Le plasmalemme est principalement composé par des glycérides (l'alcool constitutif étant le glycérol).Ex : le GPI glycosyl phosphatidyl inositol (2 alcools : glycérol et inositol). Le GPI est exclusivement situé dans le feuillet extracellulaire. Il sert de point d'ancrage à certaines protéines qui ne traversent pas la double couche (comme la phosphatase alcaline).
Les sphingolipides sont aussi nombreux. L'alcool est la sphingosine (alcool aminé). Ex : la phosphatidyl-choline (lécithine). Ce sont des molécules neutres. Elles sont toujours situées dans la couche externe. Les phosphatidyl éthanol amines, et les phosphatidyl sérines (céphalines) sont quant à elles chargées négativement et situées dans la monocouche interne.
Toute membrane biologique est polarisée. Cette polarisation électrique s'explique en partie par la composition des deux monocouches. (en tout cas c'est facile à retenir :-)). Les charges négatives se répartissent à l'intérieur, tandis que les charges positives se trouvent plutôt face externe de la membrane.
Si les lipides forment la structure de la membrane, les protéines assurent sa fonction. La plupart des protéines sont fonctionnelles et dépendent du tissu auquel appartient la cellule. Une minorité de protéines sont structurales.
On distingue deux types de protéines selon leur critère d'extraction :
- Les protéines intrinsèques / intégrées / intégrales : elles sont insolubles dans les solvants aqueux. On ne peut les extraire que si on détruit la membrane par des détergents.
- Les protéines extrinsèques / périphériques / adventives : elles sont facilement solubles dans une solution tamponnée acide ou basique dont la force ionique est relativement élevée. On n'a pas besoin de détruire la membrane pour les isoler. Ces protéines sont faiblement liées aux lipides.
Les protéines intrinsèques les plus intéressantes sont les protéines transmembranaires dont un ou plusieurs segments traversent la membranes. Ces segments sont constitués d'acides aminées hydrophobes (Isoleucine, Valine, Leucine) organisés en hélices a. D'autres protéines intrinsèques ne sont accrochées qu'à une extrémité de la membrane par des liaisons covalentes. Les protéines extrinsèques ne pénètrent pas la membrane. Elles peuvent être associées à d'autres protéines par des liaisons faibles ou aux lipides avec des liaisons ioniques.
Plus la membrane est spécialisée plus le nombre d'espèces protéiques est faible. Ceci est valable pour différentes cellules mais aussi au sein même des différentes régions du plasmalemme.
Il n'y a pas de glucides dans les membranes biologiques sauf dans le plasmalemme.
Les glucides représentent 5 à 10 % du poids sec de la membrane.
Ils sont mis en évidence par les lectines. (glycoprotéines isolées de plantes végétales). Les lectines sont très spécifiques de certains sucres. Ils servent donc d'outils spécifiques d'identification des sucres. Les lectines peuvent être couplées avec des fluorochromes.
Les glucides sont toujours liés soit à des protéines, soit à des lipides. Les glucides ne sont jamais tous seuls. Ils sont placés dans l'hémimembrane externe. C'est le cell coat.
Ils permettent la reconnaissance entre cellules. Ils sont souvent les supports antigéniques. Ex : les groupes sanguins.
Pour être accrochés aux lipides, le motif glucidique est assez simple. Ils forment les sphingolipides. On parle de cérébroside quand il n'y a qu'un ose, et de gangliosides lorsqu'il y a une chaîne osidique.
Nomenclature :
|
G => ganglioside
|
M / D => nb d'ac. sialique(s)
|
n° => 5 - nb d'oses
|
|
GM1
|
ganglioside monoacide sialique
|
4 oses
|
|
GM2
|
ganglioside monoacide sialique
|
3 oses
|
|
GD2
|
ganglioside diacide sialique
|
3 oses
|
En fonction des techniques, notre vision de la membrane a évoluée.
- Grâce au MET, Damon et Danielli mettent au point le modèle lamellaire. Il s'agit d'une structure tripartite où les lipides sont organisés sous forme de deux couches, leurs pôles hydrophobes se regardent. Les protéines auraient toutes une localisation périphérique. Ce modèle rigide ne permet pas d'expliquer comment les molécules d'eau pouvaient traverser la membrane. (L'artéfact chimique qui fausse l'observation)
- En 1972, Singer et Nicholson fondent le modèle de la mosaïque fluide, grâce à la technique de cryofracture. Au lieu de faire fixation par des molécules chimiques, on effectue une fixation par le froid en plongeant les cellules dans l'azote liquide. Ceci permet d'obtenir une image plus proche de la réalité. On sait en effet que les fractures d'échantillons congelés s'effectuent au niveau des bicouches lipidiques des membranes. On obtient donc deux hémimembranes complémentaires. C'est une de ces deux hémimembranes que l'on observe au MET. On voit alors des creux et des bosses. Les bosses représentent les protéines, tandis que les creux représentent les protéines présentent dans l'autre hémimembranes (se sont leurs empreintes). On désigne face E, l'hémimembrane exoplasmique / extracellulaire, et par face P, l'hémimembrane protoplasmique / intracellulaire.
Les protéines extrinsèques ne sont pas communes aux deux faces. Parmis les protéines intégrales on recense face E, les glycolipides associés à ... ... Face P, les protéines du cytosquelette comme l'actine. Le cholestérol est associé aux lipides.
La densité en protéines est plus importante face P ( 3000/µm vs <1500µm face E). Il y a plus de protéines du côté interne.
L'architecture moléculaire n'est pas figée.
Les lipides membranaires peuvent être animés de quatre mouvements différents : des mouvements de faible amplitude, dus à l'agitation thermique comme la flexion ou la rotation, et des mouvements de diffusion. La diffusion latérale est le déplacement des lipides au sein de la membrane (ce qui dépends aussi de la température). C'est un mouvement très rapide : 1 à 2 µm/s => en une seconde, un lipide peut faire le tour de la cellule. La chaleur et les acides gras insaturés fluidifient la membrane. Le cholestérol et les acides gras saturés la rigidifient. La diffusion transversale correspond au changement de monocouche lipidique. Elle ne dépend que de la température et nécessite l'intervention d'enzymes. On parle de flip-flop (in/out). Ce sont des flippases qui font flipper la membrane. 
Les protéines ne sont animées que de deux mouvements : la rotation et la diffusion latérale. C'est le déplacement des lipides qui en est la cause. L'asymétrie entre les deux hémimembranes est perpétuelle, il n'y a jamais de flip flop pour les protéines. La diffusion latérale est ralentie par l'attachement à des protéines cytosquelettiques (actine), par l'interaction à des éléments du cytosquelette et par l'interaction avec des protéines identiques qui se groupent en agrégats volumineux.
Il existe deux grands types d'échanges : avec ou sans déformations visibles du plasmalemme. On parle d'échanges cytotiques lorsqu'il y a déformation du plasmalemme. Ces échanges sont de grande amplitude (microorganismes, virus, bactéries, débris cellulaires). On parle d'échanges perméatifs quand il n'y a pas déformations. Ce sont des échanges à petite échelle (protéines, ions)
Il s'agit d'endocytose si une entité entre dans la cellule et d'exocytose si la cellule rejette quelque chose.
On distingue deux types selon d'endocytose selon la taille de l'élément capté. La phagocytose concerne les éléments solides de grande taille ≥1µm (ex bactérie), et entraîne la formation d'une vacuole ou phagosome de diamètre ≥ 2500Ǻ. La pinocytose concerne l'ingestion d'éléments liquides en suspension de taille ≤1µm. La vésicule formée, nommée vacuole ou vésicule, a un diamètre moyen de 1000Ǻ.
Toutes les cellules sont capables de l'effectuer.
C'est la forme non spécifique de la pinocytose. La vitesse de déplacement est strictement proportionnelle à la concentration de la substance. Une invagination localisée du plasmalemme permet la libération d'une vésicule. Cette vésicule peut fusionner avec d'autres vésicules ou fusionner avec les lysosomes. La formation de la vésicule résulte du détachement de la cupule en ampoule puis vésicule grâce à la fusion de l'hémimembrane externe avec elle-même. Il est certain qu des protéines favorisent cette étape : les protéines de fusion. (Ex : les virus utilisent les protéines de fusion pour entrer dans la cellule.
C'est une forme spécifique qui nécessite la présence de récepteurs. Elle permet à la cellule de faire entrer des molécules présentent à très faible concentration et de la concentrer dans la cellule. Ceci fait appel à la notion de couple récepteur-ligand. L'efficacité de cette forme d'endocytose est 1000 fois supérieure à celle de la pinocytose en phase liquide. L'absorption s'effectue au niveau de zones, ou puits bordés / coated pits, où une couche protéique sous jacente est présente, c'est le complexe de clatherine. Ce complexe protéique majeur est formé par des triskélions. Le triskélion est une association en trimère d'un couple de deux protéines fibreuses avec une chaîne lourde (180 kDa) et une chaîne légère (40kDa). La surface des zones bordées représente seulement 2% de la surface du plasmalemme. Une cellule compte 500 à 1000 puits. L'adaptateur à la clatherine (protéine d'assemblage) permet l'accrochage de la clatherine à l'hémimembrane interne. Les récepteurs sont soit localisés strictement au dessus des zones de clatherine, soit ils sont localisés légèrement à côté. Dans ce cas la couche de clatherine se superpose à la partie interne des récepteurs lorsque le ligand est présent.
Le cholestérol fait parti des molécules ingérées par cette pinocytose. Le cholestérol est véhiculé par des particules de faible densité : les LDL. ( Æ : 220 Ǻ, càd légèrement plus gros qu'un ribosome). Le LDL est composé de la protéine apoB (ou B 100 : PM > 500 kDa) qui est associée à des lipides et rassemble dans sa concavité 1500 molécules d'esters de cholestérol. Quand une cellule a besoin de cholestérol, elle synthétise des récepteurs à LDL. Les cellules endocrines à hormones stéroïdes en comptent beaucoup. (ex : les cellules du cortex surrénal). Les récepteurs sont des protéines transmembranaires. La partie N-ter est extracellulaire, y fait suite un site de libération pour un segment intramembranaire unique (1 hélice alpha), puis le domaine C-ter intracellulaire de liaison à la clatherine.
La première étape de fixation ne nécessite pas d'énergie, c'est juste une reconnaissance, par contre le détachement de la vésicule nécessite de l'ATP. Lorsque les vésicules bordées entrent, elles perdent leur bordure. La dépolymérisation des triskélion consomme de l'énergie. Les protéines Hsp70 favorisent cette dépolymérisation ATP dépendante. La vésicule devenue lisse fusionne avec de petites vésicules dont le pH est acide : les CURL (Compartiment de désaccouplement / uncoupling du récepteur et du ligand). Dans l'endosome tardif, le ligand se sépare de son récepteur. Il y a ensuite ségrégation ligand / récepteur. Les ligands vont se retrouver dans une partie de la vésicule. La partie contenant les récepteurs "vides" va être recyclée sur le plasmalemme tandis que la partie avec les ligands va fusionner avec le lysosome où ils seront dégradés.
Ce phénomène est assez général et n'est pas exclusif du LDL et du cholestérol. Il concerne par exemple les asiloglycoprotéines (les 2 derniers oses de la chaîne glucidique sont galactose et acide sialique). Au cours du vieillissement des protéines, l'acide sialique peut se détacher (le sucre terminal est donc le galactose). Ces protéines ne sont généralement plus fonctionnelles. Les hépatocytes du foie qui possèdent les récepteurs à galactose réabsorbent ces vieilles protéines et les dégradent.
L'hypercholestérolémie familiale (maladie autosomique dominante dont le gêne se situe sur le chromosome 19) est due à la mutation des gênes des peptides récepteurs au LDL. Le récepteur modifié possède une configuration anormale, et la fixation est mauvaise, ou il est carrément absent si la zone transmembranaire est modifiée.
La potocytose est une variante de la pinocytose par récepteur interposé où les récepteurs membranaires sont toujours fixés au GPI. (rappel : le GPI est exclusivement situé sur la monocouche externe). La couche sous jacente à la vésicule est dans ce cas la cavéoline (qui se substitue à la clatherine). La vésicule qui se forme, appelée cavéole, ne se détache pas du plasmalemme. Une fois formée la cavéole perd la couche de cavéoline. Le contenu de la cavéole s'acidifie grâce à des pompes à protons. Comme dans l'endosome tardif, le ligand se désaccouple du récepteur et traverse la membrane grâce à des perméases. Une fois le cycle terminé, la cavéole se remet en place et récupère son revêtement. La cavéole peut aussi s'isoler complètement et fusionner avec l'appareil de Golgi. (mais elles ne fusionnent jamais avec les lysosomes).
La potocytose est moins fréquent que la pinocytose classique.
C'est la voie d'entrée de la toxine du choléra. La sous-unité B se fixe sur la membrane des cellules au niveau du GM1. Une fois la toxine fixée, les deux sous-unités se dissocient et seule la sous-unité A1 pénètre dans la cellule par l'intermédiaire d'un récepteur à GPI. C'est cette dernière sous-unité A1 qui est pathologique.
La diacytose est encore une variante de pinocytose. Elle s'effectue chez les cellules épithéliales. En effet certains épithélium sont rendus étanches par des jonctions serrées entre les cellules. Pour traverser l'épithélium, les substances sont obligées de traverser les cellules. Ex : épithélium intestinal (=> entérocytes) Pour les enfants qui ingèrent le lait maternel, les anticorps du lait traversent les entérocytes par un système de récepteurs interposés. La récepteurs ne sont jamais liés à du GPI. Ils ne sont pas forcément situés dans des zones bordées.
Déroulement du processus : les particules se fixent sur les récepteurs, la vésicule se forme. Il n'y pas de phénomène de désaccouplement dans la cellule. La vésicule fusionne avec le plasmalemme de la face opposée, et alors seulement le ligand se sépare du récepteur. De la membrane basale, la vésicule va se recycler pour se remettre en place au pôle apical. Chez la mère, le phénomène inverse a lieu. Les anticorps présent dans le sang vont traverser l'épithélium de la glande mammaire par transcytose.
La phagocytose permet de capter des éléments solides. Elle est spécifique de certaines lignées cellulaires. Chez l'homme on parle de phagocytes (cellules sanguines de la lignées blanche. Ex : monocytes, macrophages, leucocytes neutrophiles). Les macrophages mangent les vieux globules rouges (hématies). Il y a dix milliards de globules rouges mangés par jour. Les macrophages peuvent phagocyter les débris cellulaires lors de réactions inflammatoires ou des organismes pathogènes => défense immunitaire passive.
Déroulement du phénomène : Ce qui est dans le milieu extracellulaire s'attache à la membrane par des récepteurs plus ou moins spécifiques. Ce phénomène n'est pas continuel mais induit. La présence de ligands déclenche la réponse. Au lieu qu'il y ait invagination du plasmalemme, il y a une évagination. Des prolongements de la cellule, les pseudopodes, englobent le corps extérieur. Les pseudopodes nécessitent des filaments d'actine. En effet, si on drogue les macrophages avec de la cytochalasine B pour détruire l'actine, les pseudopodes n'apparaissent pas. Pour dégrader le corps extérieur, le phagosome fusionne ensuite avec des Curl et des lysosomes. Les phagosomes peuvent se former sur des zones bordées ou non.
Les macromolécules destinées à être exportées sont le plus souvent enfermées dans des vésicules qui se déplacent dans la cellule jusqu'au plasmalemme : les vésicules de sécrétion. On distingue l'exocytose constitutive de l'exocytose provoquée. La migration des vésicules ne peut se faire qu'en présence d'une stimulation. Dans le cas d'exocytose provoquée, il y a des récepteurs qui fixent des ligands stimulateurs.
Par exemple, l'augmentation du Ca2+ intracellulaire déclenche la fusion des vésicules d'exocytose avec le plasmalemme. La surface du plasmalemme croît pourtant le volume cellulaire n'augmente pas. Il existe simultanément des vésicules d'endocytose pour équilibrer. Ceci a été démontrer pour les vésicules synaptiques à acétylcholine. Une protéine est présente dans la membrane des cellules synaptiques : la synaptophysine.
Les échanges perméatifs ne mettent pas en jeu de déformation visible du plasmalemme. Les matériaux traversent directement le plasmalemme. Ils ne sont jamais enfermés dans un compartiment et n'impliquent pas le cytosquelette. Certains de ces échanges consomment de l'énergie. Soit la dépense d'énergie est obligatoire et il s'agit de transport actif, soit la dépense n'est pas nécessaire et le transport est passif. On distingue aussi lorsqu'une protéine membranaire intrinsèque est indispensable au transfert.
Il s'agit d'un échange passif sans transporteur.
Pour une substance donnée, la loi qui prévaut pour entrer dans une cellule est la même que celle qui prévaut pour en sortir. Une substance donnée utilise la simple diffusion lorsque la vitesse de diffusion est linéaire avec la concentration extracellulaire. Plus une substance est hydrophobe plus l'échange est rapide. Le sens du gradient de concentration règle le sens du transport (comme pour toute diffusion). Les échanges s'arrêtent à l'équilibre. Les molécules qui participent à échange sont neutres, polaires ou non et de petite taille. Ex : urée, glycérol, benzène, éthanol, eau... Pour les molécules d'H2O, on parle d'osmose. L'eau se déplace du milieu hypotonique au milieu hypertonique. Une entrée d'eau correspond à la turgescence. Dans le cas inverse on parle de plasmolyse. Les gaz utilisent aussi la diffusion grâce aux différences de pression partielle (Ssss...)
La diffusion facilitée n'a pas besoin d'énergie mais utilise des transporteurs. Les sens du transport se fait aussi dans le sens du gradient de concentration. L'intérêt est la rapidité du phénomène. Il est mis en évidence par son caractère saturable (le nombre de transporteurs pour une substance donnée étant limité). La courbe de diffusion avec le plateau de saturation à Vmax évoque la courbe de réaction enzymatique obéissant à la loi de Mikaelis (càd dépendant du substrat) mais la molécule transportée n'est pas modifiée).
Les transporteurs qui conduisent une molécule sont appelés uniport, uniporteur, perméase ou translocase. Des transporteurs peuvent conduire 2 molécules soit simultanément dans le même sens pour les symports, soit simultanément en sens inverse pour les antiports. Les protéines transmembranaires possèdent un ou plusieurs sites de liaison de la ou les molécules à transporter. Ces protéines ont au minimum une structure tertiaire et peuvent avoir deux configurations différentes : ping et pong. Les deux états traduisent l'accessibilité de la protéine au ligand. État pong (out) : accessibilité extracellulaire, le ligand peut entrer. État ping (in) accessibilité intracellulaire, le ligand peut sortir. Le changement de conformation s'effectue de manière allostérique. Les transporteurs ne se déplacent pas dans la membrane (il n'y a pas de flip-flop). Les transporteurs sont toujours des protéines allostériques.
Dans le cas du transport du glucose, la protéine possède 12 hélices alpha et les parties N-ter et C-ter sont à l'intérieur de la cellule. On distingue Glut 1 chez les hématies et le cerveau, Glut 2 chez les cellules du foie, du pancréas, et des entérocytes, Glut 4 pour les muscles et les adipocytes.
Dans le cas du transport de l'eau, des canaux facilitent la diffusion : les aquaporines. Elles possèdent 6 hélices alpha, N-ter et C-ter sont à l'intérieur de la cellule, les segment externes sont glycolisés. La protéines fonctionne en tétramères. Un canal est formé de 4 protéines intramembranaire. Son diamètre est de 75 Ǻ (7,5 nm). Ces protéines sont fortement exprimées dans les cellules perméables à l'eau : les cellules rénales du néphron (près du glomérule). Pour les cellules perméables, les aquaporines représentent 4 à 5 % des protéines membranaires. Ces aquaporines ne sont pas spécifiques à l'eau, l'urée et le glycérol les utilisent aussi.
La diffusion accélérée concerne les molécules électriquement chargées et nécessite l'action de canaux ioniques ou tunnels. Le sens du transport d'un ion dépend de la concentration de l'ion de part et d'autre de la membrane et du potentiel électrique du plasmalemme. C'est donc le gradient électrochimique qui régit le sens du transport. On distingue deux catégories de transporteurs. Tout d'abord, les canaux voltaïques potentiels dépendants (pour Na+, K+, Ca2+, Cl-...) ; ils sont constitués d'un peptide unique à plusieurs hélices alpha pouvant se polymériser (cas du K+). Et aussi les canaux ligands dépendant ; ceux ci s'ouvrent quand une molécule agit du côté intra ou extracellulaire (exemple des neurotransmetteurs avec Ca2+ et GTP. (guanosine triphosphate). Ce sont des complexes moléculaires compliqués (récepteurs nicotiniques à acétylcholine constitué de 4 polypeptides différents).
La diffusion accélérée est un transport actif, elle nécessite donc un système de production d'énergie, la plupart du temps une enzyme s'en charge : la GTPase. Il n'y a jamais d'équilibre avec ce type d'échange car c'est un transport contre nature (comprendre dans le sens opposé au gradient de concentration).
La pompe Sodium / Potassium :
Il y a un couplage entre le flux entrant de K+ et le flux sortant de Na+. (Rappelons ici que pour une cellule vivante humaine, le Na+ est en comparativement en plus forte concentration hors de la cellule, tandis que le K+ est lui plus concentré dans le milieu intracellulaire) Il existe dans la membrane des cellule une ATPase, cette enzyme est caractéristique de la pompe Na/K. La protéine possède un site actif tourné vers l'intérieur de la cellule. Cette enzyme nécessite un environnement ionique particulier. Elle est Na/K dépendante. Le transporteur qui permet le flux ionique et le système de production d'énergie ATPase sont un seul et même système. Son inhibiteur est la ouabaïne. Cette dernière molécule bloque la pompe en prenant la place du K+. Cette protéine fonctionne comme les protéines de type ping-pong. Elle est allostérique.
Quatre étapes et des changements de conformations sont nécessaires au processus de transport.
1) État Ping (le site accessible est intracellulaire) // Fixation du Na+ intracellulaire // fixation de l'ATP => ADP +Pi, le Phosphate sert à phosphoryler le complexe // => 1er changement de conformation.
2) État Pong (le site accessible est intracellulaire) // Expulsion du Na+ fixation du K+ // Déphosphorylation => changement allostérique.
3) Pénétration du K+ retour à l'état ping.
La pompe dépense beaucoup d'énergie. 25 à 30 % des dépenses énergétiques de la cellule sont consacrées à son fonctionnement
Conséquences du contrôle des concentrations par la pompe : il permet le contrôle du volume de la cellule (grâce au jeu des pressions osmotiques). De plus il offre la possibilité de faire pénétrer sucres, oses, acides aminés. C'est un transport indirect (ou transport actif secondaire) : l'entrée de glucose ou des acides aminés n'est pas directement lié à une dépense d'énergie, mais il utilise le gradient Na/K.
Entrée du glucose, exemple de entérocytes :
Les pompes Na/K des cellules de l'épithélium intestinal ont une localisation basolatérale (en gros, en haut, en bas mais pas sur les cotés). [Na+]extérieur > [Na+]intérieur donc par diffusion le sodium passe de la lumière intestinale dans la cellule. Simultanément le glucose entre aussi. Il y a cotransport de type symport. Le glucose ressort au niveau basal de la cellule à l'aide d'une diffusion facilitée (ici seul le glucose passe). Le cotransporteur symport est un homodimère . Chaque sous unité possède un site Na et un site glucose. La liaison du sodium précède la liaison du glucose. Cette protéine a 12 hélices alpha. Parmis les 664 acides aminés qui composent la protéine, l'aa 28 est essentiel. Si l'acide aspartique est remplacé par l'asparagine, le transporteur n'est plus fonctionnel.
Transport des acides aminés
A la partie apicale des cellules, se trouve un cotransporteur