Généralités :
A Historique.
En 1680 Anton Van Leeuwenhoeck a fait les premières observations en microscopie optique avec un grossissement de 300 fois : c'est la première observation des micro-organismes, appelés à l'époque «animalcules ».
Au 19e siècle, Pasteur donne une nouvelle notion des micro-organismes, en précisant leur rôle, et il donne une nouvelle idée de la vaccination.
I Notions.
Les travaux de Pasteur sur les ferments démontrent absurdité de l'idée de génération spontanée. Il démontra ceci grâce à cette expérience:
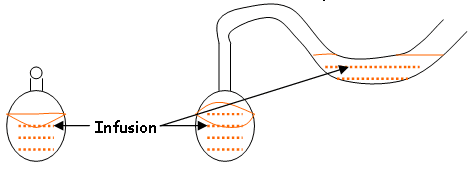
Si l'on isole l'infusion de l'air ambiant, il n'y a pas de contamination : le milieu est stérile. Les premiers travaux sur les infusions de foin de Pasteur, ont été repris par Tyndall : il faisait subir une pasteurisation mais observait toujours un développement. Il a donc mis au point une méthode à plusieurs chauffages : la tyndallisation. Cette technique est utilisée contre les bacillus.
II Rôle.
Ce fut l'avènement de la microbiologie médicale. Celle-ci a été permise par Koch et par Pasteur. Ces deux chercheurs ont trouvé un protocole sur les agents causants des maladies. Les premiers essais ont été réalisés sur les moutons avec la maladie du charbon (bacillus anthracis). Le bacille de Koch a pour nom systématique : Mycobactérium tuberculosis.
Le but de la microbiologie est de trouver les micro-organismes responsables des maladies. Brefeld a eu l’idée d’un milieu de culture solide et Pétri a eu l’idée de la boite du même nom.
La vaccination est une mise en contact de formes atténuées d’agents microbiens. La première vaccination est celle contre la rage qui a été réalisée vers 1880.
1 La période moderne.
a 1918 à 1928 et de 1928 à 1952.
C’est à ce moment là que l’on a de nouvelles identifications de maladies et de micro-organismes. En 1928, Flemming travail sur Staphylococcus sp. mais sa boite est contaminée par un pénicillium : il y a inhibition de l’activité de Staphylococcus : c’est le premier antibiotique.
b Travail sur Escherichia coli.
Depuis 20 ans, c’est l’avènement du génie génétique. On a assisté à l’apparition de techniques de clonage, d’addition de gènes… le plus souvent grâce à des plasmides de bactéries. Ces travaux ont généralement été réalisés sur Escherichia coli.
B Présentation.
I Généralités.
On utilise (ou on a utilisé) plusieurs mots pour parler des micro-organismes : microbe, animal pur, germes, ferments, protistes. Ce sont des organismes unicellulaires, pluricellulaires ou coloniaux à l’intérieur desquels n’a pas lieu de différenciation cellulaire. On observe toutefois des cas spéciaux comme chez les virus, les champignons et des algues. Toutefois, de nombreuses bactéries ne peuvent se développer seules : elles forment donc des colonies (on parle alors de myxobactéries). Certains signaux cellulaires sont assez puissants pour faire se regrouper une colonie bactérienne. Chez les cyanobactéries, on observe des cellules spécialisées comme les hétérocystes.
II Caractéristiques.
Les micro-organismes sont partout, nombreux car très diversifiés sur un plan taxonomique (on parle de pléiomorphisme). Ces organismes occupent tous les milieux en quantité importante. Ils ont en plus de grandes capacités métaboliques.
III Eléments de classification.
On a trois domaines :
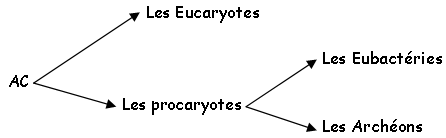
• Les eucaryotes sont présents dans quatre règnes : chez les protistes, les végétaux, les mycètes et chez les animaux.
• Chez les eubactéries ont trouve par exemple Escherichia coli.
• Chez les archéons, on trouve des organismes de type extrémophiles, thermoacidophiles, dans les dorsales océaniques. Ils ont une structure interne qui est intermédiaire entre celle des procaryotes et des eucaryotes.
IV La théorie des endosymbiontes.
La principale différence entre les procaryotes et les eucaryotes est la présence d’organites chez les cellules eucaryotes. On note aussi la présence de stérols spécifiques aux eucaryotes. On trouve des glycérophospholipides membranaires chez les eubactéries et les eucaryotes qui sont toutefois différents. Ces molécules sont aussi présentent chez les archéons mais leur composition est intermédiaire entre les deux autres.
Chez les eucaryotes, on trouve des diester d’acide gras. Chez les archéons on trouve des diéther d’alcool polyisoprénoïdes qui sont des précurseurs des stérols.
La partie commune :
• Les ribosomes : chez les eucaryotes ils ont de type 80S et de type 70S chez les procaryotes. Ils ont le siège d’action de la streptomycine.
• La membrane plasmique bactérienne n’est pas qu’une membrane d’échanges comme chez les eucaryotes, mais aussi une membrane de synthèses.
• Les cyanobactéries possèdent des thylacoïdes comme les végétaux (mais pas de chloroplastes).
• L’ADN : chez les procaryotes, il est sous forme d’un brin circulaire (un seul chromosome). Cet ADN n’est pas associé à des histones. La duplication est réalisée au moment de la division cellulaire.
• Le brassage génétique : il se réalise pendant la fusion des deux gamètes chez les eucaryotes. Ce processus n’existe pas chez les procaryotes : dans ce cas, la cellule mère va donner deux cellules filles identiques entre elles et à la mère. Les rares changements observés ont lieu quand ce micro-organisme réalise des échanges de brin d’ADN avec le milieu extérieur.
• La taille : elle peut aller de 10Å pour les virus jusqu’à 100µm pour les protozoaires.
• Les différentes formes :
- Les archéons : On trouve par exemple Méthanobactérium qui assure la réaction CO2?CH4.
- Les eubactéries : elles sont divisées en deux sous-groupes selon la constitution de leur paroi. On trouve les gram+ et les gram-.
- Les eucaryotes : on les trouve dans trois règnes distincts : les animaux, les végétaux et les mycètes (ou Fungi ou champignons).
Parmi le groupe des mycètes, on prend l’exemple d’une levure comme saccharomyces cerevisae. Comme espèce de moisissure coenocytique, on trouve Pénicillium roquefortii (protiste).
Parmi les protozoaires, on trouve Plasmodium.
Chez les algues, on peut trouver chlorella, goniolax (algue rouge), dinophysis, les diatomées (base de la nourriture des baleines).
C Ecologie microbienne.
Vers la fin du 19ème et au début du 20ème siècle, Winogradsky et Beijeriwck travaillent sur les bactéries, ayant plus un intérêt sur l’aspect fonctionnel que sur l’aspect taxonomique.
I Les milieux.
1 Les eaux.
Les eaux contiennent une grande quantité de bactéries en suspension 10^4 bactéries de micro-plancton par mL dans l’océan et 10^11/mL dans les égouts. On trouve de 10² à 10^5 algues microphytes par mL, de 100 à 1000 champignons par mL et quelques milliers de prédateurs comme les paramécies : cet ensemble de micro-organismes forme la boucle microbienne. On remarque que les micro-organismes océaniques peuvent faire précipiter le CO2.
Les substances nutritives : dans les milieux de culture, ces quantités de substances sont exprimées en g/L alors qu’ailleurs, ces quantités sont exprimées en mg (voir µg) par L. Quand il y a trop de nutriments, on assiste alors à une eutrophisation du milieu qui va entraîner un dysfonctionnement et une surproduction (surpopulation).
Tous les types micro-organismes existent.
2 Le sol et les sédiments.
Dans les sols, on trouve peu de microphytes mais beaucoup de bactéries (10^7/g) de moisissures (10^5/g) et d’actinomycètes (10^6/g). Ces organismes sont des décomposeurs qui permettent une reminéralisation des sols. Dans un sol, il y a un phénomène de stratification qui entraîne différentes teneurs dans les éléments selon la strate. La respiration anaérobie sur le CO2 est appelée méthanogenèse.
3 L’atmosphère.
On observe une compartimentation à travers laquelle sont disséminés les micro-organismes par émission de spores. La présence de ces organismes est fonction des ressources présentes.
II Animaux et végétaux comme compartiments naturels
On trouve trois types de micro-organismes :
- Les pathogènes : ils déclenchent des pathologies (maladies)
- La microflore commensale : c’est, par exemple, notre flore intestinale, avec Escherichia coli.
- Les symbiontes : ils ont une capacité de relation complète (végétaux ou animaux avec des bactéries) pour accomplir les fonctions qu’ils ne peuvent faire qu’à deux.
Exemple : la microflore des ruminants.
On trouve deux chambres de cultures. La première chambre (le rumen) sert à la culture des bactéries micro-cellulolytique (qui digèrent la cellulose). Cette flore est ensuite digérée dans la seconde chambre (le tractus). Les Méthanobactérium du rumen produisent du méthane (CH4).
III Les cycles.
1 Le carbone.
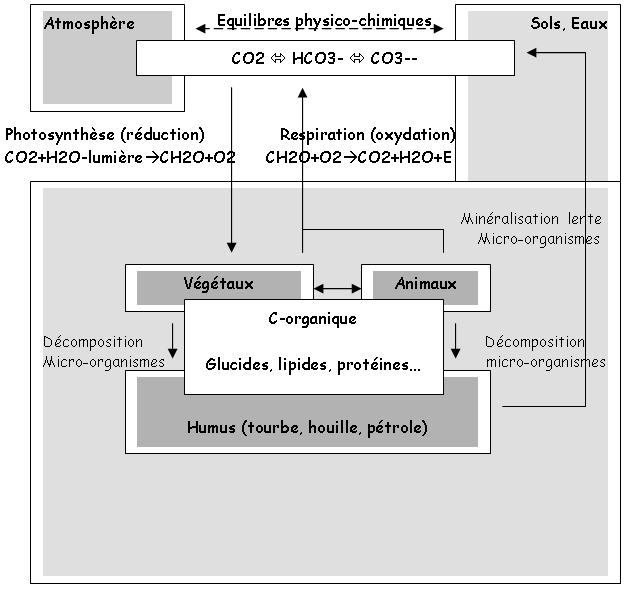
Ce cycle décrit les principales formes chimiques, les compartiments où elles sont engagées, les flux entre compartiments et les équilibres qui en résultent. Le recyclage du carbone concerne 60 à 90% de la matière organique produite.
2 L’azote.
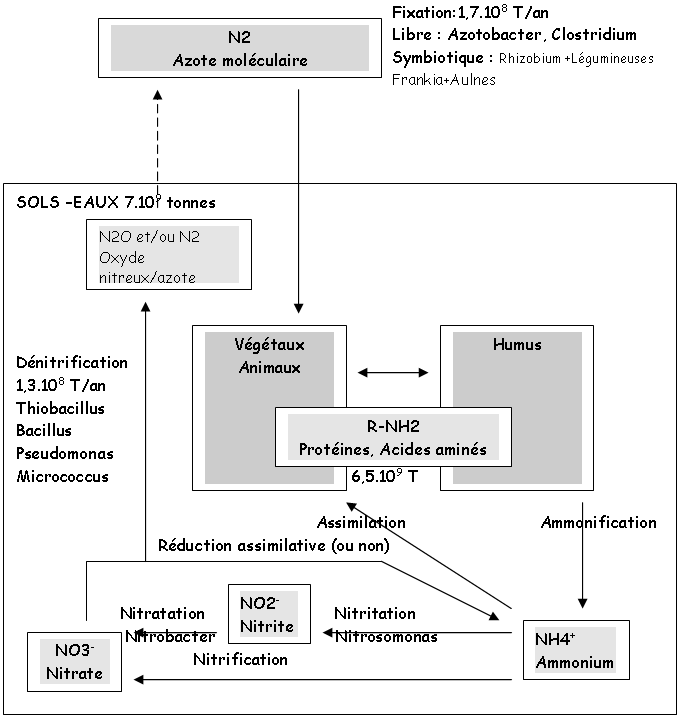
L’azote doit être fixé pour pouvoir être incorporé dans le cycle de la vie.
N2+6(e-)+6H+ 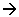 2NH3.
2NH3.
Cette réaction est réalisée par une enzyme de type nitrogénase (chez les procaryotes). Azotobacter et les hétérocystes des cyanobactéries sont des fixateurs libres de l’azote.
Les symbioses réalisées par le rhizobium créent des nodosités sur les racines des plantes.
La nitrification : les nitrifiants sont chimio-autotrophes (C venant de CO2, N venant de NH4+ ou de NO2-). NH4+ + 3/2O2 NO2- + 2H+ + H2O. Exemple de cette réaction avec Nitrobacter ou Nitrosomonas.
La dénitrification : dans ce cas, l’azote sert d’accepteur final d’électrons à la place de l’oxygène. NO3- N2O ou N2.
Chapitre 1 : Structure des micro-organismes.
I Les bactéries.
A Généralités.
Les micro-organismes sont les plus petites unités biologiques fonctionnelles. Leur taille est comprise entre 0,01 et 10 µm.
Les différentes formes existantes :
• Les coques : ils sont sphériques. Les différents assemblages de coques proviennent de la division cellulaire.
• Les bâtonnets (bacilles) : ils sont droits (coli bacille), incurvés comme les vibrions (exemple : Vibrio cholerae). On peut aussi trouver des formes irrégulières et enflées aux extrémités (exemple : mycobacter, corybacter). Une autre forme possible est la forme coccobacille (exemple : Serratia marcescens).
• Les hélicoïdaux : on trouve les spirochètes qui pourraient être à l’origine des mouvements des premières cellules eucaryotes
• Les formes mycelliennes : ce sont des formes filamenteuses, comme les Actinomycètes. Les myxobacters ont des organes de fructification qui sont des ensembles de bactéries de forme similaires.
B Structure.
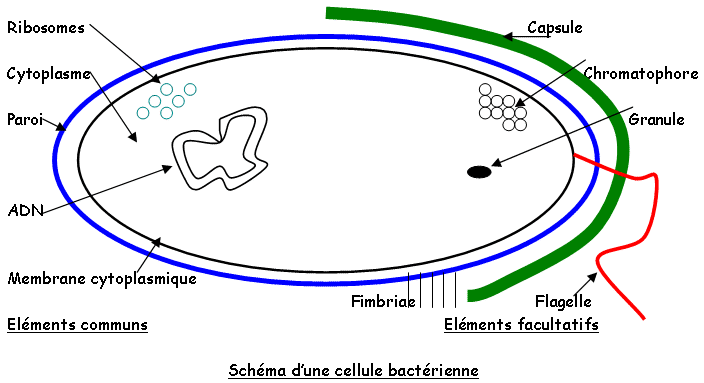
Les micro-organismes sont étudiés par fractionnement, par immunocytologie, ou au microscope électronique à balayage (MEB). On a un schéma type de bactéries avec des composants types comme les ribosomes, le cytoplasme, la paroi, la membrane cytoplasmique et l’ADN. On peut aussi trouver des composants facultatifs comme les granules, les réserves, les chromatophores, des fimbriae, des flagelles et des capsules.
1 La membrane plasmique.
Cette membrane a une structure classique, en double feuillet, mais moins rigide que les membranes eucaryotes car elles ne possèdent pas de stérols (sauf les mycoplasmes).
a Composition.
Cette membrane est composée de 30 à 40% de lipides (dont les glycérophospholipides) et de 60 à 70% de protéines.
b Rôle et fonction.
C’est une barrière semi-perméable qui permet le transport passif de certaines molécules grâce à des protéines canales.
Les protéines membranaires sont des enzymes permettant des biosynthèses de lipides bactériens ou de peptidoglycanes.
La membrane peut avoir un rôle respiratoire grâce à la présence de cytochromes dans cette membrane (un peu comme une mitochondrie).
2 Le cytoplasme.
Il contient 80% d’eau avec un pH compris entre 7 et 7,2 où sont dissous des sucres, des ions, des acides aminés… On y trouve aussi des inclusions de granules de polyphosphates ou de polyhydroxybutirate, des ribosomes associés à l’ARNm sous forme de polysomes (1000/cellule).
3 Le nucléoïde.
C’est le génome bactérien (ou chromosome bactérien). Il peut exister sous plusieurs copies en même temps. On assiste au phénomène d’amitose : il n’y a pas de synchronisation entre la division de l’ADN et celle de la cellule.
a Organisation.
Ce nucléoïde n’est pas isolé du cytoplasme par une membrane. Il est constitué par un double brin circulaire (refermé) d’ADN : c’est un assemblage en double hélice de deux chaînes antiparallèles et complémentaires de nucléotides. (A et G sont puriques alors que C et T sont pyrimidiques).

L’ADN bactérien est sans histones et est super-enroulé. Très souvent, ce dernier est ancré en un ou plusieurs points de la membrane plasmique. Par exemple, chez Escherichia coli, on trouve 5 millions de paires de base où 4300 gènes sont identifiés (60% du total). Cet ADN mesure 1mm quand il est déroulé et représente 10% du volume cellulaire.
b Rôle.
C’est le support de l’hérédité. Ce nucléoïde permet de lire le génome, de le transcrire en ADN (afin de synthétiser des protéines). Il subit aussi la réplication pour assurer la descendance.
c Biosynthèse de nucléoïdes.
La réplication est la synthèse d’un nouveau génome : c’est un mécanisme semi-conservateur et bidirectionnel.
Il apparaît une fourche de réplication grâce à une DNA-polymérase. Il y a phosphodiestérification entre l’amorce d’ADN et ce qui est lu dans le sens 3’5’.
On parle de brin sens quand il y a réplication dans le même sens que celui de l’ouverture de la fourche de réplication.
Les fragments synthétisés sur le brin anti-sens sont reliés par une ADN-ligase.
Les deux brins sont synthétisés en même temps dans les deux sens.
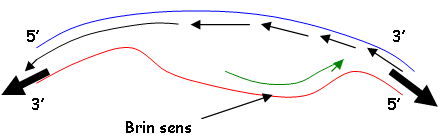
L’ADN-gyrase permet de désenrouler au point d’origine et suit le mouvement des fourches. Cette enzyme est inhibée la norobiocine (antibiotique).
d Les plasmides.
* Définition : ce sont des molécules d’ADN double brin, circulaire, qui, extrachromosomique, ne constituent pas le génome bactérien. Ils ont une réplication autonome (un pouvoir infectieux), une petite taille et codent pour une information génétique non-indispensable. Ils peuvent infecter des bactéries ou être échangés entre elles.
Ces plasmides ont été découverts en 1952 sur Shigella dysenteriae.
Shigella résistante + E. coli saine Shigella résistante
E. coli résistante
E. coli saine
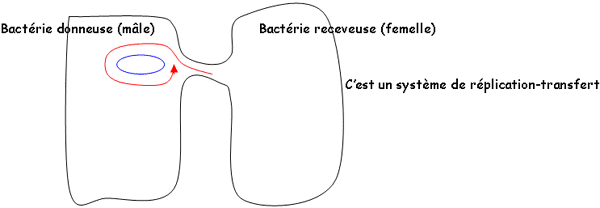
*Réplication et transfert : elle se fait de façon autonome selon le même processus que celui du nucléoïde. Ils ont la même vitesse de réplication : on peut donc avoir plusieurs plasmides en même temps dans une même cellule. Le transfert des plasmides se fait par conjugaison :
* Propriétés : les plasmides sont des unités codantes. Ils donnent à la bactérie :
- la possibilité de synthèses spéciales
- une résistance à des antibiotiques : comme des enzymes qui dégradent les antibiotiques (par exemple : la 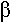 -lactamase qui résiste aux pénicillines.
-lactamase qui résiste aux pénicillines.
- Une pathogénicité. Chez E. coli, il y a synthèse d’entérotoxines qui provoquent des maladies. E. coli peut aussi gagner l’aptitude à se fixer sur une membrane.
- Un pouvoir infectieux
- Le plasmide F donne la possibilité de recombinaison génétique : il code pour la synthèse de pili sexuels pendant la conjugaison.
e La paroi.
C’est la plus externe, c’est elle définit la cellule procaryotique. De plus, celle-ci sert à la classification des micro-organismes. Elle représente 20% de la masse sèche. Elle a un rôle majeur dans la résistance à la pression osmotique et aux déformations. Cette coloration (ou non-coloration) est révélatrice d’une différence structurale de la paroi.
Organisation :
La coloration de Gram (en 1844). C’est une étape préliminaire pour la reconnaissance d’une bactérie. Cette coloration est réalisée en présence d’iode, puis on lave à l’alcool : on trouve alors deux cas distincts : - les bactéries sont décolorées : ce sont des gram-
- les bactéries gardent la coloration : ce sont les gram+.
La paroi des gram+ est 1000 fois plus grande que celle des gram-. La paroi des G+ est généralement composée de peptidoglycane et d’acide teïchoïque. Ce dernier représente 50% du poids de la paroi structurée.
La paroi des G- est composée, sur la membrane externe, de lipoprotéines, de LipoPolySaccharides et de trimères : c’est un réseau lâche.
Remarque : Les mycoplasmes ne sont ni l’un ni l’autre : ils n’ont pas de paroi. Les archéons ont une structure un peu différente.
Composition globale :
Dans les parois de G-, on trouve plus d’acides aminés et de lipides.
• Les macromolécules spécifiques :
- L’acide teïchoïque : il est composé, soit de ribitol (CH2OH-(CHOH)3-CH2OH), soit de glycérol. Ils sont substitués avec du glucose et/ou de l’alanine. Cet acide peut avoir un rôle de reconnaissance antigénique (sérotypie). Ces acides sont ancrés, soit dans la membrane, soit dans le peptidoglycane (où ils sont pariétaux). Ils servent aussi à relier la membrane et la paroi. Plus le réseau formé est dense, plus la paroi est rigide.
- Le LPS : chez les G-, il constitue les feuillets externes de la paroi externe de cette bactérie. Ce LPS est composé en trois parties
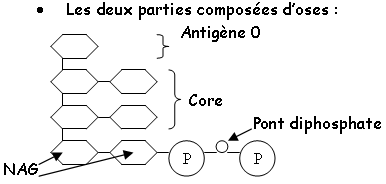
• Le rôle du LPS : il a le même rôle antigénique que l’acide teïchoïque d’ancrage et de structuration de la membrane externe.
- Les protéines de la membrane externe des G- :
On trouve deux grands types : les lipoprotéines qui sont des polypeptides qui lient la membrane externe au peptidoglycane et les protéines matricielles qui traversent la membrane externe et qui peuvent avoir un rôle dans le transport (porines) ou dans la réception des phages.
- Le peptidoglycane : (mureïne, glycocalix). Il a un haut poids moléculaire et est spécifique aux eubactéries. Il assure un rôle structurant (résistance et pression). Il donne donc la forme cellulaire et empêche la lyse par un milieu hypotonique.
- Les constituants :
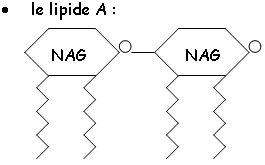
• Les osamines : ils sont composés de NAcétylGlucosAmine ou d’Acide NAcétylMuramique : c’est le composant de la paroi bactérienne. UDP-NAG + PEP + NADPH UDP-NAM + NADP. Cette réaction est inhibée par la phosphonomycine. Chez les archéobactéries, il n’y a pas de muréine mais de la pseudo-muréine (pas de NAM mais du N.AcétylOsaminUronique).
• Les acides aminés :
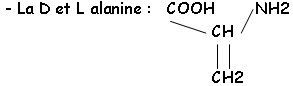
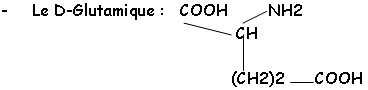

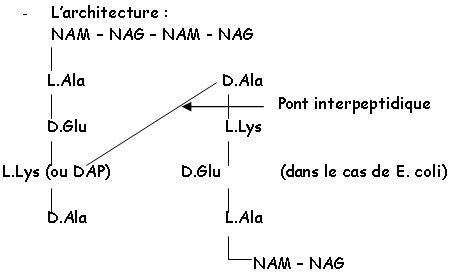
Cette chaîne est réticulée grâce au NAM et à son substitue en 3’ : il y a relation avec la L.Ala, la L.Lys ou le DAP, le D.Glu et la D.Ala.
Chez Staphylococcus aureus, on a L.Lys – (Gly)5 – D.Ala : c’est un maillage lâche qui lui donne sa forme sphérique.
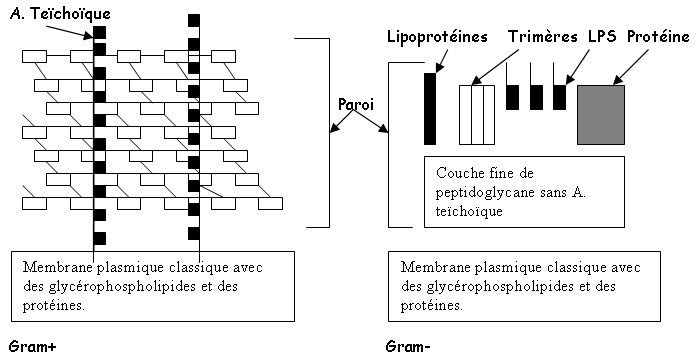

Remarque : les mycoplasmes sont des parasites intracellulaires, donc dans un milieu isotonique. L’enveloppe comprend la paroi et la membrane.
Le rôle de la paroi : c’est une barrière (active chez les G- pour les transports). Le peptidoglycane assure la forme cellulaire. La couche la plus externe a des propriétés :
- antigéniques : des particules induisent la production d’anticorps : on obtient donc une définition de sérotypie bactérienne (grâce à l’acide teïchoïque chez g+, au LPS chez les g-.
- de fixation des phages (bactériophages = virus à bactéries). La recombinaison génétique à lieu grâce aux bactériophages. N’importe lequel de ces phages ne peut pas infecter n’importe quelle bactérie. Chez E. coli, le T4 se fixe sur le LPS, le  se fixe sur la protéine qui transporte le maltose. Chez bacillus suptilis, le
se fixe sur la protéine qui transporte le maltose. Chez bacillus suptilis, le 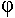 29 se fixe sur l’acide teïchoïque.
29 se fixe sur l’acide teïchoïque.
f Flagelles et pili.
* Les flagelles : ils sont facultatifs, de nature protéique (la flagelline), sont des unités sphériques en hélice et sont ancrés dans la membrane plasmique. Ils per(mettent les déplacements microbiens : ils ont donc un rôle de chimiotaxie. Ils ont aussi des propriétés antigéniques : ils permettent de fixer les phages de type PBS1 sur Bacillus suptilis. On trouve différent type d’insertion des flagelles :
Le type d’insertion des flagelles peut être utilisé pour des reconnaissances bactériennes.
* Pili (ou fimbriae) : ce sont des éléments facultatifs de la cellule g-, ils sont de nature protéique comme les flagelles, mais en général plus courts. Ils procurent à la bactérie qui les portent, une capacité d’adhérence (souvent associée à la virulence de la bactérie). Ils ont un rôle dans la reconnaissance entre cellules donneuses et receveuses pendant la conjugaison. Ils sont aussi le site de fixation des phages comme le M13.
g La capsule.
C’est l’enveloppe supérieure la plus externe, souvent polysaccharidique (composée souvent d’acide hyaluronique). Cette capsule est associée à une virulence comme chez Klebsiella pneumoniae. Cette enveloppe entraîne un phénomène d’adhérence, mais elle masque aussi les sites antigéniques, et augmente la taille apparente de la bactérie pour résister à la phagocytose. Elle permet aussi la résistance à des conditions externes défavorables.
h Les endospores.
La sporulation est un phénomène induit par une carence nutritive. La germination nécessite une activation (choc thermique, forte variation de pH). La sporulation est une mise en l’abri du génome (le record de longévité est de 7500 ans) face aux conditions défavorables du milieu (dessiccation : perte d’eau du milieu).
Parfois, la formation des spores est associée à une libération de toxines. Par exemple, Bacillus thuregensis sécrète un cristal particulièrement herbicide.
La structure des spores :
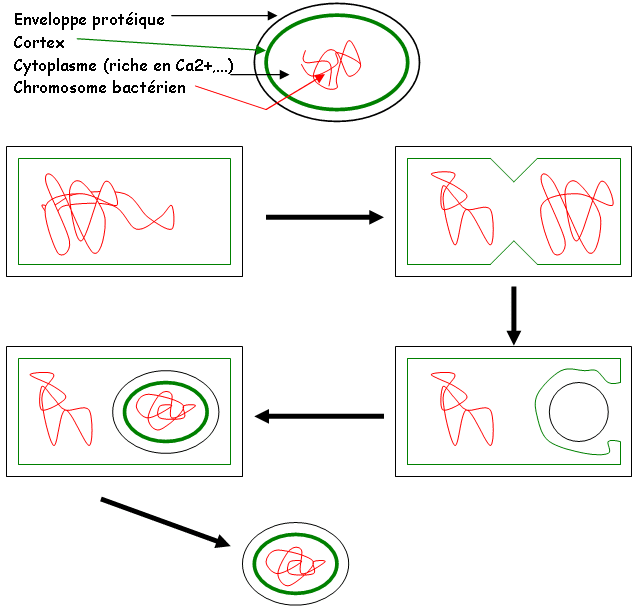
C Classification.
La classification est réalisée par des
- Critères morphologiques (structuraux)
- Critères métaboliques (type respiratoire, condition de vie)
- Pili, endospores
C’est une approche phénotypique.
On peut aussi avoir une approche moléculaire pour se baser sur le génotype.
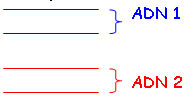
On va chauffer l’ADN pour le dénaturer. Ensuite, on observe si les ADN se recombinent ou non :

On a aussi une approche en pourcentage de G-C : ce pourcentage donne la température de dénaturation.
On peut aussi étudier les séquences d’ADN robosomiaux. Ils ont les propriétés d’être indispensables à la vie et de subir des variations pendant l’évolution. Ils permettent d’obtenir une distance génotypique. Exemple :

Chapitre 2 : Les Mycètes.
I Introduction.
A la place de mycètes, on peut entendre parler de mycota ou de fungi. Ces espèces (les mycètes) forment un règne du vivant. Ils sont caractérisés par des formes comme les moisissures, les mycètes à carpophore volumineux (champignons).
On a sur Terre, à peu près 100.000 espèces de mycètes. Une centaine est pathogène pour les hommes et les animaux et un autre millier est pathogène envers les végétaux.
Ces mycètes sont des micro-organismes eucaryotes : leur masse de cytoplasme est pluri ou mono-nucléée. Ce cytoplasme est contenu dans un système de tubes (les hyphes) ramifiés (le mycélium). Les hyphes sont rigidifiés par une paroi.
Dans ce règne, on observe la présence de structures spécialisées, comme le carpophore qui est un objet sexuel.
Les cycles biologiques sont différents selon les espèces de mycètes (haplophasique, diplophasique, haplodiplophasique).
Tailles et formes :
- On trouve des espèces unicellulaires uninucléées, dont le diamètre est compris entre 5 et 10µm, comme saccharomyces cerevisae.
- D’autres espèces sont filamenteuses (mycelliennes).
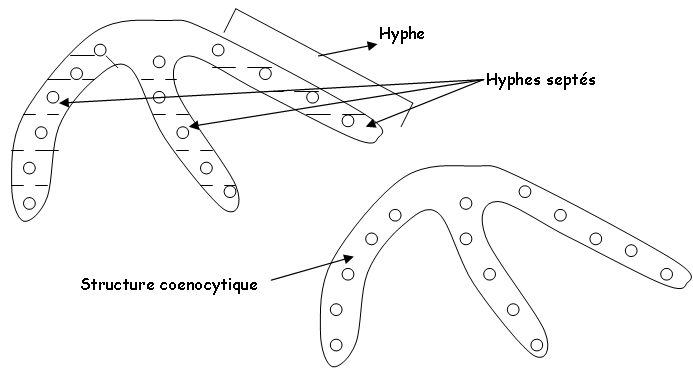
Remarques :
- Les actinomycètes ont la même organisation mais avec les propriétés des procaryotes qu’ils sont.
- Un Armillaria bulbosa s’étendait sur 15ha, pesait 100 tonnes et avait plus de 1500 ans.
- dimorphique : c’est une espèce qui est sous forme filamenteuse à basse température (milieu solide) et sous forme unicellulaire à 37°C en milieu liquide (exemple : Candida albicans).
II Reproduction.
A Reproduction asexuée.
1 Bourgeonnement.
C’est le type de reproduction des levures du type de Saccharomyces (sauf pour les pseudo saccharomyces).
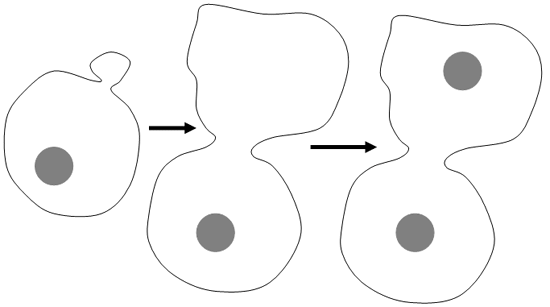
2 Spores et conidiospores.
Ils sont fabriqués par des hyphes spécialisés (les conidies) d’un mycélium aérien. Les conidiospores vont donner la couleur à l’hyphe.
B La reproduction sexuée.
On a quatre groupes distincts de reproduction sexuée. La classification est basée sur la structure dicaryote : fusion de deux mycéliums de sexe opposé (hétérothallisme).
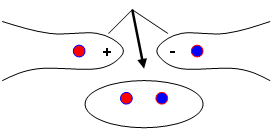
Les mycéliums sont haploïdes et leur fusion donnera donc un individu diploïde.
1 Les zygomycètes.
Exemple : Rhizopus stolonifère.
La reproduction sexuée a la forme d’un zygosporange. La fécondation est une caryogamie (réunion des deux noyaux sans passage par les gamètes). Cette fécondation est suivie d’une méiose qui permet le passage du zygosporange au sporange.
2 Les ascomycètes.
On trouve dans ce groupe les levures (exemple : Saccharomyces cerevisae), des champignons (Tuber melanosporum = la truffe) ainsi que de nombreux pathogènes.
Un mycélium dicaryote va être isolé. Il a ensuite caryogamie. Cette dernière va permettre d’arriver à 4 ou 8 spores contenues dans un asque.
3 Les basidiomycètes.
On trouve dans cette catégorie des champignons classiques (à gros carpophore) comme le cèpe (Boletus edulie).
La basidie est une structure qui est le siège de la caryogamie. Celle-ci provient d’un mycélium dicaryote qui résulte de la fusion hétérothallique de deux mycéliums de sexe opposé.
Remarque : pour en savoir plus, consulter le chapitre 6 du cours de Biologie Végétale.
III Mode de vie.
Les mycètes sont des organismes hétérotrophes. Ils se nourrissent par absorption de composés solubles (osmotrophes). Ils sont équipés d’un système digestif extracellulaire : il y a libération d’exo-enzymes (avec la cellulose) : ce sont de remarquables décomposeurs.
Il existe trois principaux modes de vie : les saprophytes, les symbiontes et les parasites.
• Les moisissures sont les premières formes de vie de mycètes apparues. Les mycètes ont une croissance rapide et se multiplient souvent de façon asexuée (surtout chez les saprophytes et les parasites)
• Les levures : elles sont dans les milieux aquatiques.
• Les lychens sont des symbioses à bénéfices réciproques entre un mycète et une algue.
• Les mycorhizes : ce sont des associations de racines de cormophytes et de mycélium.
IV Structure.
Les mycètes ont des noyaux individualisés, leur cytoplasme est riche en mitochondries, golgi et réticulum endoplasmique. Leur membrane plasmique contient des stérols (spécifiques aux eucaryotes).
A Les lipides membranaires.

La plupart des antifongiques est dirigée contre la synthèse de ces ergostérols. L’Allylamine empêche la formation de la molécule, l’Immidazole et le Triazole sont dirigés contre une isomérase.
B La paroi.
On trouve trois grands types de polymères (planche 15). On ne trouve pas de ponts interpeptidiques comme dans le peptidoglycane des bactéries, mais un réseau de fibrilles.
1 La chitine.
C’est un polymère de N-AcétylGlucosAmine. C’est aussi un analogue de la cellulose. La chitine n’est pas spécifique des champignons (on la retrouve chez les insectes (les fourmis)). Sa synthèse est inhibée par la Polyoxine.
2 Le glucane et le manane.
Ce sont des composés avec de longues chaînes en 1-6 (de glucose ?) pour le manane et avec des ramifications en 1-3 pour le glucane.
La chitine, le glucane et le manane sont les trois composés qui assurent la rigidité des parois de mycètes. On note aussi, dans ces parois, la présence de glycoprotéines et de nombreux ponts di-sulfures.
Chapitre 3 : Les Virus.
I Introduction.
A Définition.
Ce sont des acides nucléiques infectieux, encapsulés dans une enveloppe protéique. Ils possèdent soit de l’ADN, soit de l’ARN. Ils sont obligatoirement parasites et présentent de nombreux antigènes.
B Historique.
Ils ont été découverts par Pasteur.
Iwanowsky, en 1892, met en évidence les différences entre virus et bactéries.
En 1935, Stanley isole et cultive des virus.
C Caractéristiques.
• Leur taille maximale est de 50nm.
• Ce sont des agents infectieux destinés à parasiter.
• Ils détournent la machinerie cellulaire à leur profit (ils peuvent parasiter n’importe quel être vivant).
• Un virus possède un seul type d’acide nucléique (ADN ou ARN).
• Il n’y que quelques cas où l’information génétique est portée par l’ARN.
II Structure.
L’assemblage d’un acide nucléique et d’une capside protéique forme la nucléocapside. On a parfois l’existence d’une membrane lipidique qui va former l’enveloppe.
A L’acide nucléique (ADN ou ARN).
Il est double ou simple brin. L’ADN est souvent sous forme linéaire et est composé de 1000 à 2500 nucléotides. Au maximum, on a 50 gènes.
Le brin sens (+) peut servir d’ARNm alors que le brin anti-sens ne le peut pas.
B Les protéines.
On trouve essentiellement des enzymes : la transcriptase inverse dans le cas du VIH, l’ADN-polymérase quand l‘information est sur l’ARN, des lysosomes qui permettent l’hydrolyse du peptidoglycane (chez les bactériophages).
C La capside.
On trouve des capsomères et des nucléocapsides.
D L’enveloppe.
C’est un élément facultatif résultant du bourgeonnement de la cellule hôte. Elle est très souvent constituée des constituants (« comme c’est logique ») de la cellule infectée et des spicules de la cellule virale.
Les spicules sont des toxines de nature protéiques. On trouve par exemple, l’hématogluttinine et la neuramidase.
E La classification.
Elle repose sur la nature de l’ADN ou de l’ARN, du type de symétrie, de la présence ou non de l’enveloppe.
III Propriétés.
Les virus sont des parasites obligatoires. De plus, ils ont des propriétés immunologiques grâce à l’enveloppe associée aux spicules et grâce à la capside.
Dans le type Influenza, l’ARN du génome est fragmenté : on obtient donc un génome différent grâce à des mutations, dans ce cas, très faciles.
IV Les cycles viraux.
Tous les cycles sont composés de trois étapes successives :
1. Phase de pénétration, de décapsidation et d’absorption
2. Synthèse.
3. Maturation (assemblage de nouveaux virions) et libération.
Quand il y a intégration du génome viral au génome bactérien (à la cellule hôte, en réalité), on assiste à une lysogénie.
A Les bactériophages (deux types).
On trouve des phages tempérés qui ont un cycle lysogénique (série ) et des phages virulents qui eux, ont un cycle lytique.
• Le cycle lytique :
les phages de type T (planche 18, figure1 à gauche : ils ont une grande spécificité des fonctions pour les interactions. Ils effectuent une reconnaissance grâce à des fibres codales.
- Le lysozyme : c’est une enzyme qui hydrolyse la membrane des G-.
- La gaine virale : par un phénomène d’hydratation, cette gaine se contracte pour injecter le génome viral dans la cellule hôte.
- Le génome, une fois à l’intérieur, se cyclise puis détruit le génome bactérien
- Il y a ensuite transcription de l’ADN phagique et synthèse d’ARNm. Cet ARNm sert à la fabrication d’enzyme de réplication pour des protéines de futures particules phagiques.
- Après la maturation, il y a assemblage des éléments de la queue du phage. Ces nouveaux virus seront relargués dans le milieu grâce au lysozyme.
• Le cycle lysogénique :
- Il a interaction de l’ADN phagique et du génome bactérien. Ce phénomène a lieu, le plus souvent, quand il y complémentarité de séquences entre les deux ADN.
- Il y a alors intégration du génome viral dans le génome bactérien : on obtient un prophage.
- On assiste ensuite à la dissémination du prophage à l’état latent dans les bactéries filles.
- Le passage de l’état latent du prophage à un état actif se fait par une stimulation (ou induction).
Remarque : Le prophage serait à l’origine de Clostridium botalinum.
B Les adénovirus.
Ce sont des virus de cellules eucaryotes (planche 18, figure 2).
- L’absorption : elle se fait sur un site de reconnaissance membranaire.
- La pénétration : elle se fait par pinocytose. La particule virale arrive dans le cytoplasme où à lieu la décapsidation (migration de l’ADN du virus vers le noyau).
- Dans le noyau, on a des synthèses précoces : on a des transcriptions et traductions pour des enzymes de réplication et des synthèses d’ARNm pour les synthèses tardives (dans le cytoplasme)
- Les protéines du capside sont synthétisées.
- Les particules virales sont alors maturées.
- On a ensuite la lyse cellulaire et la libération des nouveaux virus.
C Les rétrovirus (exemple du VIH).
Ce sont des virus à ARN qui s’attaquent aux eucaryotes. Ils sont basés sur l’existence d’une enzyme : la transcriptase-inverse qui permet la synthèse d’ADN à partir d’ARN (planche 19).
- Il y a plusieurs absorptions par pinocytose
- Décapsidation de l’ARN et synthèse d’ADN dans le cytoplasme grâce à la transcriptase-inverse (présente dans le rétrovirus). On a alors une synthèse d’ADN double brin.
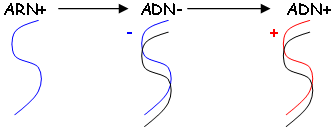
Dans le noyau, une enzyme intégrase, va intégrer le génome viral au génome cellulaire. La différence avec le phage est qu’il y a toujours intégration du génome viral au génome cellulaire. Il y a réplication et transcription d’ADN viral pour fabriquer de l’ARNm et l’ARN de support génétique.
Dans le cytoplasme, il y a synthèse d’intégrases, de transcriptases-inverses… La libération des nouveaux virus se fait par bourgeonnement. Quand il n’y a pas d’activité virale, on dit que le virus est éteint.
Remarque : les virus à oncogènes provoquent des cancers.
Chapitre 4 : Les Antibiotiques.
I Introduction.
A Historique.
A la fin du 19ème siècle, on savait que la quinine permettait la lutte contre le paludisme.
• En 1909, Paul Ehrlich donna l’idée de «magic bullet » qui permettrait d’atteindre la partie infectante mais pas l’hôte.
• En 1928, Flemming découvre la pénicilline.
• En 1939, R. Dubois trouve l’antibiotique «gramicidine » à partir d’un Bacillus trouvé dans le sol.
• En 1944, Waksman isole, à partir d’un actinomycète, la streptomycine (contre la tuberculose).
B Définition.
Un antibiotique, au sens strict, est une substance naturelle produite par un micro-organisme dont l’action est de limiter la croissance (action bactériostatique) ou de tuer (bactéricide).
On a :
• Des antibiotiques d’extraction : ils sont souvent bactériostatiques
• Des antibiotiques de synthèse grâce à l’intervention de l’homme pour leur fabrication.
• Des agents anti-infectieux : ils sont «anti-tout ».
Les antibiotiques sont différents des antiseptiques car ils ont des actions ciblées sur un mécanisme spécifique (métabolisme cellulaire).
Le spectre d’activité : il définit les différents micro-organismes touchés par un antibiotique. Le spectre est lié au mode d’action du mécanisme considéré.
II Les familles d’antibiotiques (planche 13).
Les antibiotiques ont des origines variées :
- Bacillus : ce sont des g+, à faible G-C% (à endospores) qui fabriquent des polypeptides.
- Actinomycètes : ils sont à l’origine de 70% des antibiotiques et 60% pour la seule espèce de Streptomyces.
- Céphalosporium : de sont des organismes eucaryotes à l’origine de 10% des antibiotiques.
On a quatre types de spectre :
• anti g+
• anti g-
• antifongiques
• spectre large
III Mécanisme et action.
A Au niveau du fonctionnement cellulaire.
On a cinq grands types (classés dans l’ordre d’importance croissante) :
- Inhibition de la synthèse des métabolites essentiels.
- Inhibition de la transcription et/ou de la réplication de l’ADN (novobiocine, rifampicine).
- Dénaturation de la membrane plasmique (polypeptides).
- Inhibition des synthèses protéiques (action bactériostatique) (streptomycine).
- Inhibition de la synthèse de la paroi.
B Cible moléculaire.
Le mécanisme d’action d’un antibiotique a une cible particulière et précise.
• Les -lactamines : elles inhibent l’enzyme responsable de la synthèse du peptidoglycane, sur la transpeptidase. Cette enzyme est inhibée par la pénicilline qui se fixe sur le site actif et prend la place du peptidoglycane.
• La streptomycine (aminoside) : cet antibiotique agit la synthèse des protéines. Elle se fixe sur la petite sous-unité du ribosome. Au niveau de la formation du complexe, elle perturbe la fixation de l’ARNm, d’où, des erreurs de lecture.
IV Résistances.
La résistance est un mécanisme acquit par une bactérie qui était sensible à un antibiotique et qui y devient résistante. Les principaux mécanismes d’acquisition d’une résistance sont dus à des mutations génétiques ou à l’infection par un plasmide.
Les deux formes de résistance à la streptomycine :
- Il peut y avoir une mutation du gène codant pour la petite sous-unité 30S du ribosome. La streptomycine n’a plus de relation avec cette sous-unité. Si cette dernière reste fonctionnelle, on obtient une forme de résistance.
- Le gène Sp de plasmide qui code pour l’adényl-transférase va greffer un radical adényl sur streptomycine qui devient alors trop grosse pour pouvoir se fixer.
La résistance à la pénicilline est souvent due à des processus d’infection. Il y a synthèse, par la bactérie, de -lactamase (enzyme) qui inactiver la -lactamine qui devient alors inopérante.
On a alors, trois manifestations possibles :
- Il y a non-reconnaissance de la cible.
- La molécule d’antibiotique voit sa forme être modifiée.
- La cellule perd sa perméabilité pour l’antibiotique (pour les tétracyclines).
Remarque : le gène Tet (sur un plasmide) fait ressortir l’antibiotique.
La résistance grâce aux plasmides est la résistance la plus répandue et la plus facile à répandre. Le plasmide R permet la propagation des épidémies nosocomiales (les infections en milieu hospitalier).
Chapitre 5 : Métabolisme et Génétique.
I Métabolisme.
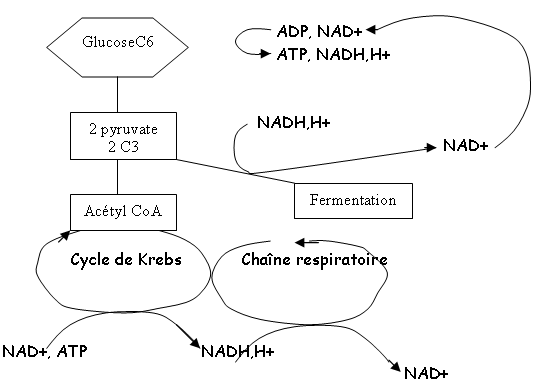
Le métabolisme est la somme des activités servant à fabriquer de la cellule (anabolisme) et des activités servant à produire de l’énergie (catabolisme).
Les fonctions du catabolisme des chimio-hétérotrophes forment le métabolisme oxydatif. Grâce aux réactions rédox, on a la récupération de l’énergie résultant de l’oxydation d’un composé (formation d’ATP).
A Le métabolisme glucidique.A
1 Le glucose.
a La formation du pyruvate.
• La glycolyse
1 C6 2 C3
Bilan : 2 ATP + 2 NADH,H+ (le NADH,H+ est présent pour récupérer les électrons au cours des réactions rédox.
• La voie du KDPG
C’est une voie alterne à la glycolyse.
1 C6 2 C3
Bilan net : 2 NADH,H+ + 1 ATP.
• La voie des pentoses-phosphates.
Cette voie est complémentaire à la glycolyse. Elle a un rôle de dégradation des sucres en C5 pour former des C4.
Bilan : 3 C6 3 C5 + 3 CO2 + 9 NADH,H+.
b Le devenir du pyruvate.
On a cinq possibilités :
- fermentation : le pyruvate devient un substrat (il peut être réduit en acide lactique). La respiration se combine avec le coenzyme A qui passe dans le cycle de Krebs.
- La respiration aérobie : il y a production d’ATP et accumulation de NADH,H+. Ces deux composés vont aller alimenter les chaînes respiratoires (dans les mitochondries ou dans les membranes bactériennes) et régénérer du NAD+. NADH,H+ NAD+ ; O2 H2O ; 1 C6 6CO2. On a un bilan total de 38 ATP pour les procaryotes et de 36 ATP pour les eucaryotes (2 ATP sont utilisés pour traverser la paroi mitochondriale). Et le NAD+ est régénéré.
- La respiration anaérobie : l’accepteur final peut être différent type de molécules. On peut trouver des nitrates (NO3- N2), des sulfates (SO4^2- H2S), CO2 CH4. Ce sont des chaînes incomplètes : le bilan donne toujours 6CO2 mais moins de 38 ATP et moins de NAD.
- Les fermentations : leur but est de régénérer le NAD+. Elles ont le même bilan que la glycolyse.
1 C6 2 C3 Acide lactique
+ 2 ATP
+ 2 NADH,H+ 2 NAD+
B Les Polysaccharides.
La filière générale permet de dégrader les oses en glucose. Si on obtient un autre composé, ce sera du fructose qui rentrera plus loin dans le cycle.
La cellulose : les enzymes cellulases sont fabriquées par des micro-organismes de type Actinomycètes, Bacillus, Vibrio, Cytophage.
C Le catabolisme protéique.
On trouve les micro-organismes responsables de ce catabolisme dans les putréfactions et les gangrènes. Il y a hydrolyse des protéines pour former des amines volatiles qui donneront les mauvaises odeurs. Il y a ensuite une phase d’ammonisation : les acides aminés donnent des acides organiques quelconques et de l’ammoniac.
D Le catabolisme des lipides.
Ils sont dégradés en acétylCoA qui va rejoindre le cycle de Krebs.
• Nocardia permet la dégradation des pneus.
• Certaines bactéries stimulées vont être capables de dégrader les nappes de pétrole.
II Génétique.
C’est la recombinaison et les transferts génétiques. Ces deux phénomènes ont un rôle de reproduction sexuée chez les micro-organismes qui n’en ont pas naturellement. Les recombinaisons ne touchent que les secteurs homologues.
A Les mutations.
Une mutation est un changement spontané. Il y a une mutation pour 10^6 gènes répliqués. On obtient alors une bactérie mutante toutes les 10^7 bactéries formées. Parmi les facteurs mutagènes, on trouve les UV, les radiations…
B Les recombinaisons (parasexualité).
Les recombinaisons ont lieu grâce à trois mécanismes (peut-être quatre) et entraînent la formation d’un mérozygote (diploïde partiel)
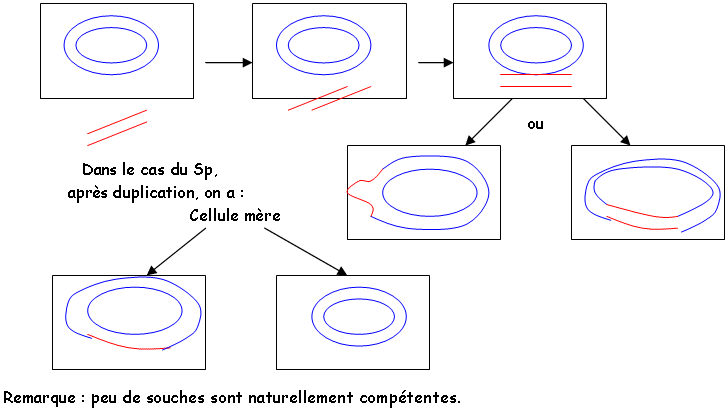
1 La transformation (l’expérience de Griffith).
On travaille à partir de Streptococcus pneumoniae sous deux souches : virulente (SpE) et non-virulente (SpN).
Si on chauffe SpE et qu’on l’injecte dans une souris, la souris ne meurt pas. Si l’on réalise un broyat de SpE avec des SpN et qu’on injecte celui-ci dans la souris, elle meurt.
En 1944, Avery, McLeod, McCarthy découvrent la solution à ce problème : l’ADN.
Une bactérie intacte associée à un fragment d’une autre entraîne une transformation. Ce phénomène demande une «compétence » de la cellule réceptrice. Cette compétence s’acquière en fin de phase de multiplication de la cellule (faculté de perméabilité).
On a trois étapes : compétence, synthèse d’ADNliase, synthèse d’ADNhydrolase.
2 La transduction.
Ce phénomène est en relation avec des virus.
a La transduction généralisée.
Un phage arrive, injecte son ADN sans hydrolyser le génome bactérien : on a alors un virus avec ADN bactérien.
b La transduction spécialisée.
Ce phénomène est réalisé par un phage tempéré. Il y a excision de l’ADN du phage du génome bactérien. Ce sont toujours les mêmes gènes qui sont concernés.
3 La conjugaison.
C’est un échange d’ADN par contact entre bactérie réceptrice et donatrice. L’ADN concerné est souvent le plasmide F. On parle pendant la conjugaison, de bactérie + (mâle) et de bactérie – (femelle). L’adhésion est permise grâce aux F-pili ou pili sexuels.Le canal de conjugaison est encore mal connu. L’ADN échangé est de l’ADN monobrin.

Remarque : le cas des mutants. On parle de HFr (High Frequency recombinaison). Dans ces cas, un épisome est intégré dans le génome bactérien. Il faudrait de 90 à 100 minutes pour échanger tout un brin. En général, seul un morceau de brin est échangé.

